Máte zapnutý náhled celé osnovy, zpět na běžné zobrazení.
Načítání a prohlížení osnovy může být v závislosti na množství obsahu pomalejší.
Genetika
1 Úvod - Genetika
2 Genetická informace
2.4 Testovací otázky
2.1 Chromosomy
2.2 Nukleové kyseliny
- Molární obsah adeninu se rovná molárnímu obsahu thyminu:
A = T; T = A
- Molární obsah guaninu se rovná molárnímu obsahu cytosinu:
G = C; C = G
- Obecně je v každé dvoušroubovici DNA obsah molekul purinových bází rovný obsahu molekul pyrimidínových bází:
(A + G) = (C + T)
Typ nukleové kyseliny |
Stavební složky | ||
Pentosa |
Dusíkaté bázy |
Fosfodiesterická vazba | |
DNA |
β-D-2-deoxyribosa |
adenin, thymin, cytosin, guanin |
estericky vázaný zbytek H3PO4 |
RNA |
β -D-ribosa |
adenin, uracyl, cytosin, guanin |
estericky vázaný zbytek H3PO4 |
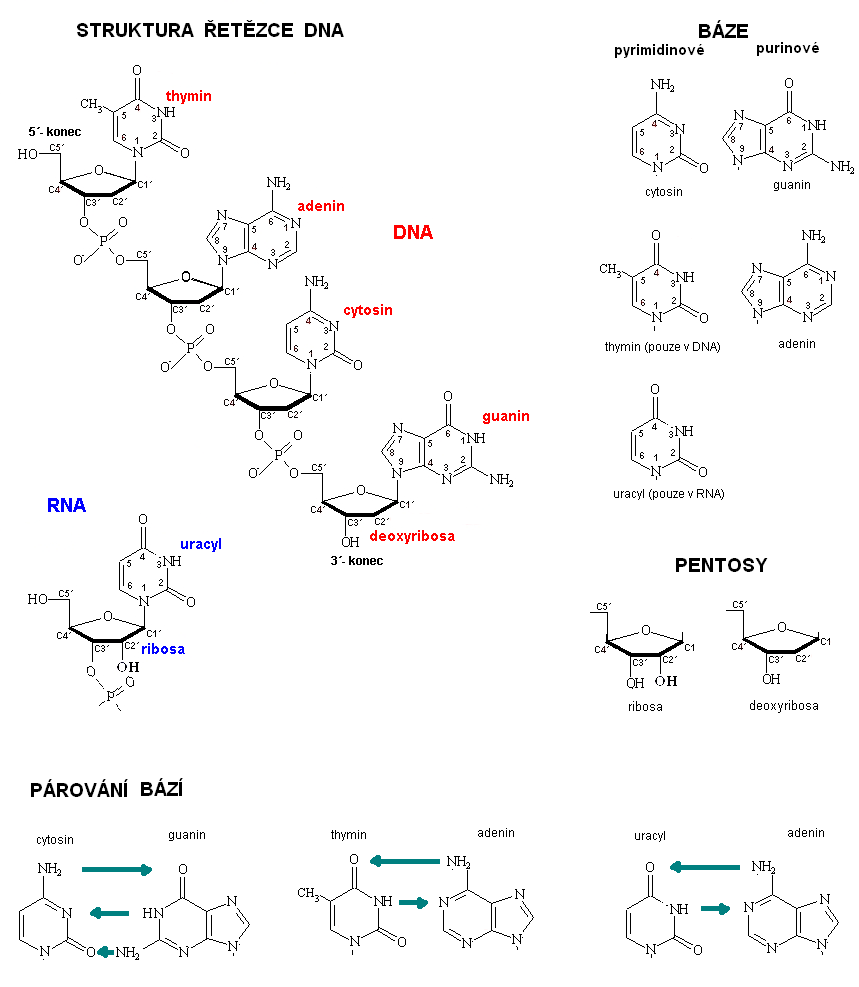
2.3 Koncepce genu
3 Zpracování genetické informace
3.5 Testovací otázky
3.1 Replikace
V průběhu replikace se řetězce rozpletou a DNA polymeráza syntetizuje oba řetězce jedním směrem prostorově a zároveň ve směru 5 '- 3' současně pro oba řetězce:
3.2 Transkripce
3.3 Translace
Báze |
U |
C |
A |
G | ||||
U |
UUU |
fenylalanin |
UCU |
serin |
UAU |
tyrosin |
UGU |
cystein |
UUC |
fenylalanin |
UCC |
serin |
UAC |
tyrosin |
UGC |
cystein | |
UUA |
leucin |
UCA |
serin |
UAA |
stop |
UGA |
stop | |
UUG |
leucin |
UCG |
serin |
UAG |
stop |
UGG |
tryptofan | |
C |
CUU |
leucin |
CCU |
prolin |
CAU |
histidin |
CGU |
arginin |
CUC |
leucin |
CCC |
prolin |
CAC |
histidin |
CGC |
arginin | |
CUA |
leucin |
CCA |
prolin |
CAA |
glutamin |
CGA |
arginin | |
CUG |
leucin |
CCG |
prolin |
CAG |
glutamin |
CGG |
arginin | |
A |
AUU |
isoleucin |
ACU |
threonin |
AAU |
asparagin |
AGU |
serin |
AUC |
isoleucin |
ACC |
threonin |
AAC |
asparagin |
AGC |
serin | |
AUA |
isoleucin |
ACA |
threonin |
AAA |
lysin |
AGA |
arginin | |
AUG |
methionin |
ACG |
threonin |
AAG |
lysin |
AGG |
arginin | |
G |
GUU |
valin |
GCU |
alanin |
GAU |
kys. asparagová |
GGU |
glycin |
GUC |
valin |
GCC |
alanin |
GAC |
GGC |
glycin | ||
GUA |
valin |
GCA |
alanin |
GAA |
kys. glutamová |
GGA |
glycin | |
GUG |
valin |
GCG |
alanin |
GAG |
GGG |
glycin | ||
3.4 Regulace genové exprese
4 Vztah genetické informace k rozmnožování
Rozmnožování a tvorba potomstva jsou základní charakteristikou živých organismů. V případě pohlavně se rozmnožujících eukaryotních organismů, mezi které patří i člověk, se musí zohlednit jejich tzv. diploidní stav. Znamená to, že v jádře buňky se nachází 23 párů chromosomů, dohromady tedy 46. Chromosomový pár tvoří typově stejnými chromosomy, které však mají různý původ. Každý pochází od jiného rodiče. Schematicky se tato skutečnost označuje symbolem "2n".
Z uvedených skutečností vyplývá, že v lidském organismu se musí využívat dva typy rozmnožování buněk. Prvním typem se zajistí, aby vzniklo potomstvo muže a ženy splynutím jejich pohlavních buněk. Druhým typem je třeba zaručit rozmnožení buněk, specifických pro tkáně a orgány. Výsledkem správného průběhu obou typů dělení je normálně vyvinutý organismus člověka.
V literatuře se jednotlivým typům rozmnožování buněk říká "dělení" buněk. Myslí se tím fakt, že během svého rozmnožování se mateřská buňka rozdělí na několik dceřiných buněk. Pokud má být rozmnožování mateřské buňky úspěšné, dceřiné buňky by měly být stejně životaschopné jako jejich "matka". Z toho vyplývá nutnost, aby zdědili všechny buněčné komponenty a informace o jejich struktuře, které jsou pro buňku životně důležité. Patří sem mitochondrie, které jsou centrem dýchacích procesů. Pro buňku jsou nepostradatelné a bez nich nemůže existovat. Dále je třeba jmenovat Golgiho aparát, endoplazmatické retikulum, které je centrem proteosyntézy probíhající na ribozomech. Nakonec nelze zapomenout ani na chromozomy, které se nacházejí v jádře a nesou genetickou informaci buňky.
Pokud se buňka chce dělit, musí dostatečně "narůst", aby zajistila dostatek stavebních a strukturálních komponentů pro všechny dceřiné buňky. Rozdělením na dceřiné buňky současně buňka mateřská zaniká.
Můžeme tedy shrnout, že rozmnožování buněk je většinou cyklický děj, který začíná vznikem dceřiné buňky rozdělením její "matky", pokračuje růstem vzniklé buňky a končí jejím dělením na další potomstvo. Tento cyklus se nazývá životní cyklus buňky. Z formálního hlediska se období mezi vznikem buňky z její mateřské předchůdkyně a jejím zánikem dělením nazývá interfáze. Interfáze se dělí na tři časové úseky: G1-fázi, S-fázi a G2-fázi.
G1-fáze se uskutečňuje jako první po vzniku nové buňky. Je typická intenzivní metabolickou aktivitou, charakteristickou pro růst. Dochází během ní k syntéze potřebných enzymů a dalších bílkovin, které jsou nezbytné pro tvorbu jednotlivých strukturních částí buňky.
S-fáze je charakteristická tím, že se během ní syntetizuje DNA. Buňka sice má "svou" DNA, kterou zdědila, ale na to, aby mohla dát šanci vzniknout minimálně dvěma dceřiným buňkám, nutně potřebuje vytvořit co nejpřesnější kopii své DNA. Musí vytvořit kopie všech 46 chromozomů, které se nacházejí v jejím jádře. Nově kopie chromozomů se však během rozdělení mateřské buňky na dceřiné nenacházejí samostatně, ale jsou spojeny se svým "originálem". Pod mikroskopem lze proto vidět typické struktury chromozomů, které jsou jedinečné pouze pro časový úsek dělení buňky. Mají charakteristický tvar připomínající písmeno X. Pozornému pozorovateli neunikne, že i po syntéze kopií chromozomů zůstává přítomen diploidní stav (2n). Abychom však novou situaci odlišili, označujeme ji symbolem 4C. Jinými slovy, z původních dvou kopií určitého chromozomového páru vzniknou další dvě nové, spolu jich je tedy 4 (proto "4C"), které však nejsou samostatné (proto nadále platí "2n", označení diploidních stavu).
G2-fáze je typická syntézou buněčných stavebních složek, potřebných pro nadcházející rozmnožování buňky. Patří sem např. mikrotubuly. Po této fázi může nastat dělení buňky na své dceřiné potomstvo. Všimněte si důležité slůvko "může". Pro organismus je nezbytné regulovat počet buněk jednotlivých tkání a orgánů. Pokud je jich dostatečný počet, pak nově vzniklé buňky se již většinou dále nedělí, ale nahrazují funkčně opotřebované předchůdkyně. V takovém případě mluvíme o speciální fázi života buňky, tzv. "G0" fáze. Je charakteristická dostavbou a dozráním - diferenciací do podoby, ve které buňka vykonává svou roli v organismu. Po jejím opotřebení buňka zaniká, nastává buněčná smrt. Ta však není (s výjimkou např. poškození a traumata) neregulovaná. Naopak, organismus se snaží využít veškerý použitelný materiál zanikající buňky. Mluvíme o apoptóze, tedy o regulované buněčné smrti.
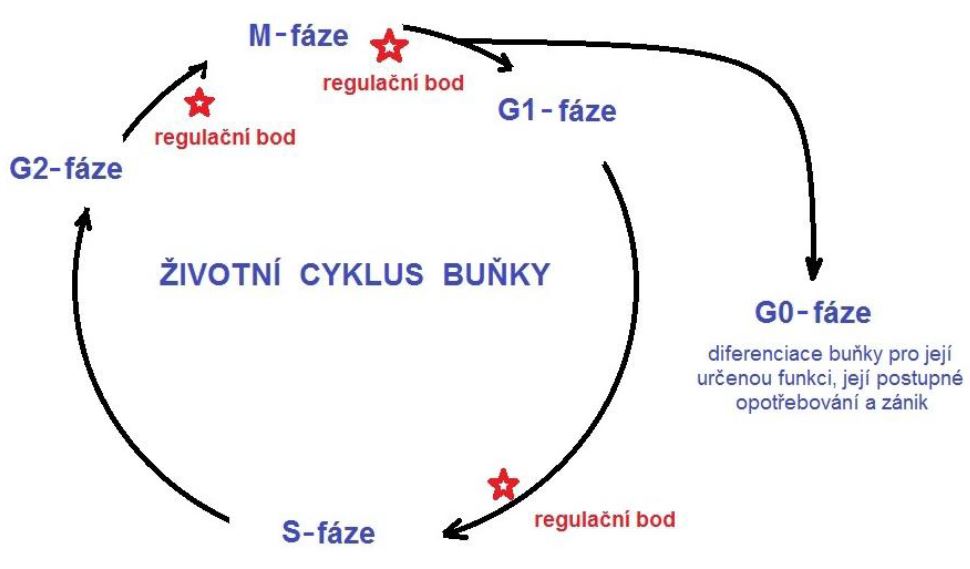
Obrázek 2 - Životní cyklus buňky
Ani sled jednotlivých životních fází není automatický, ale je regulován. Regulátory jsou speciální informační proteiny (tzv. cykliny) a jejich receptory, které mají rovněž enzymatickou funkci. Nazývají se cyklin-dependentní kinázy. Při poruše několikanásobné úrovně regulace rozmnožování buňky může nastat nekontrolované dělení buněk, které je typické pro nádorová onemocnění.
Na životním cyklu buňky vidíme, že převážnou část jejího života tvoří interfáze. Časově kratší rozsah většinou zabírá rozdělení buňky. Během dělení buňky většinou zaniká jaderná membrána. Je to proto, aby spiralizované chromozomy mohly volně nasedat na vlákna dělícího aparátu, který je rozdělí k opačným stranám buňky. Mluvíme proto o tzv. nepřímém dělení buňky. Patří sem mitóza, čili nepřímé somatické dělení buňky a meióza, která je nepřímým redukčním dělením. Obě dělení jsou podrobně vysvětleny v kapitole 4.1. Kromě nich lze výjimečně u některých živých systémů pozorovat rovněž přímé dělení buněk, tzv. amitózu, při kterém jaderná membrána nezaniká. Jádro se podélně prodlouží a rozdělí na dvě části, které jsou základem jader dvou nových dceřiných buněk. V druhém sledu se rozdělí cytoplazmatická membrána, čím se ukončí proces dělení. Amitóza bývá vzácná, častěji jde o atypicky průběh mitózy, tzv. endomitózu.
4.1 Mitosa a meiosa eukaryontních buněk
Profáze je počáteční fází mitózy, během které dochází ke spiralizaci chromosomů, proto zpravidla trvá ze všech fází nejdéle. Díky své spiralizaci jsou v této fázi chromosomy pozorovatelné pod světelným mikroskopem. Postupně zaniká jaderná membrána a vzniká dělící vřeténko složené z mikrotubulů.
V metafázi se spiralizované chromosomy přesunou do centrální části buňky (tzv. ekvatoriální roviny buňky). V optickém mikroskopu tak lze pozorovat jejich typické hvězdicovité uspořádání uprostřed buňky, které se nazývá monaster. Chromosomy se spojují s mikrotubuly dělícího vřeténka v místě své centromery. Následně dojde k oddělení obou kopií chromosomu tím, že se centromera rozpadne.
Anafáze je typická přesunem obou polovin genetického materiálu k opačným pólům buňky. Pod světelným mikroskopem je tuto fázi možné rozpoznat právě podle tzv. diasteru, hvězdicovitého uspořádání chromosomů na opačných pólech dělící se buňky.
Telofáze je závěrečnou fází mitózy. Chromosomy se seřadí kolem pólů dělícího vřeténka. Dochází k jejich postupné despiralizaci a začíná se tvořit kolem nich jaderná membrána. Zároveň dochází k rozdělení celého vnitřního prostoru mateřské buňky - cytokineze.
Výsledkem mitózy je vznik dvou dceřiných buněk, které jsou geneticky identické se svou mateřskou buňkou (obrázek 3).
Meióza není na rozdíl od mitózy natolik běžná v lidském organismu i organismech ostatních rostlin i živočichů vůbec. Jejím úkolem není produkovat identické buňky, ale napomáhat v maximalizaci variability daného druhu. Proto úzce souvisí s pohlavním rozmnožováním a tvorbou pohlavních buněk - gamet. Gamety mají pouze polovinu genetické informace (jsou haploidní, "n") v porovnání s rodiči (jsou diploidní, "2n"), kteří je tvoří ve svých pohlavních orgánech (vaječnících nebo varlatech).
Pokud se druh rozmnožuje nepohlavně, je tvořen populací geneticky téměř zcela identických jedinců. V případě nepříznivého vnějšího vlivu (změny ekosystému, patogen) je tak ohrožena celá populace, protože na nepříznivý podnět reaguje uniformně. Pokud však uvažujeme o pohlavním rozmnožování, situace se mění.
Pohlavní rozmnožování vnáší do populace důležitou vlastnost - variabilitu. Pokud máme např. muže, ženu a jejich společné dítě, z genetického hlediska se jedná o tři různé jedince. Rodiče jsou vzájemně odlišní ve své genetické výbavě. Dítě má s každým z rodičů společnou pouze polovinu své genetické informace. Představuje však novou kombinaci u původních genů, která vznikla spojením poloviny genetické informace muže a poloviny genetické informace ženy. V případě jakéhokoli vnějšího vlivu na tuto rodinu (nepříznivé prostředí, patogen) tak musíme uvažovat o rozdílných reakcích třech různých genotypů: otce, matky i dítěte. Vidíme, že variability je dosaženo právě pohlavním rozmnožováním, které vyžaduje pro vznik potomstva dvou jedinců s různým pohlavím. Meióza se proto vyskytuje v pohlavních orgánech a zajišťuje vznik pohlavních buněk (gamet), které musí mít haploidní počet chromosomů proto, aby jejich splynutím vznikl diploidní jedinec.
Meióza se skládá ze dvou po sobě jdoucích dělení: První z nich se nazývá heterotypické dělení a druhé homeotypické dělení. Heterotypické dělení má čtyři fáze: profázi I, metafázi I, anafázi I a telofázi I.
Profáze I: Na začátku meiózy lze pozorovat tzv. bivalenty, stejné chromosomy, které tvoří homologický pár. Po postupné spiralizaci jsou uspořádány vedle sebe. Kromě spiralizace chromosomů se však v této fázi probíhá i výměna nesesterských chromatid mezi oběma chromosomy, které tvoří homologický pár (obrázek 3). Nazývá se rekombinace (angl. crossing-over). Jejím úkolem je zvýšit variabilitu vzájemnou kombinací těch genů, které jsou na jednom chromosomu a nedají se volně kombinovat. Podobně jako v případě mitózy i při meióze představuje tato fáze relativně dlouhé časové období, které je členěno na pět stádií: leptotenní, zygotenní, pachytenní, diplotenní a diakinézi. Jaderná membrána mateřské buňky postupně zaniká.
Metafáze I: Po úplné spiralizaci se chromosomy soustřeďují v ekvatoriální oblasti buňky a spojují se s mikrotubuly dělícího vřeténka.
Anafáze I: K opačným pólům buňky se přesouvají celé chromosomy, tvořené dvěma kopiemi (chromatidy), které jsou spojené centromerou. V tom se anafáze I liší od anafáze mitózy, v níž se centromery rozpadají. Výsledkem anafáze I je snížení počtu chromosomů v budoucích dceřiných buňkách na polovinu (obrázek 3). Každý z obou chromosomů má 50% pravděpodobnost, že se dostane do nově vytvářených gamet. Mluvíme o "rozchodu" chromosomů do dceřiných buněk, o jejich segregaci.
Telofáze I: Dochází k částečné despiralizaci chromosomů, které jsou již soustředěny ve dvou základech haploidních jader. Systém je připraven na homeotypické dělení.
Na obrázku 3 můžeme vidět, že výsledkem heterotypického meiotického dělení jsou dvě dceřiné buňky, které sice mají z dvojice chromosomů pouze jeden (n), ten je ovšem tvořen dvěma kopiemi - chromatidami (2C). Proto musí následovat ještě jedno dělení, které zajistí, aby se obě kopie od sebe oddělily a rozešly do dvou dceřiných buněk. To je úkolem homeotypického dělení, které se tak nazývá proto, že je svým průběhem velmi podobné mitóze. Má čtyři fáze: Profázi II, metafázi II, anafázi II a telofázi II. Jeho výsledkem je rozdělení dvou dceřiných buněk, které pocházejí z heterotypického dělení na výsledné čtyři dceřiné buňky, které mají poloviční (haploidní, „n“) počet chromosomů v jejich jediné kopii (1C, obrázek 3).
Přestože obě pohlaví využívají ve svých pohlavních orgánech meiózu, mezi spermatogenezí muže a oogenezí ženy existují významné rozdíly. Spermatogeneze trvá od puberty po celý život muže a tvoří se miliony spermií. Oogeneze začíná již v prenatálním období ženy a pokračuje od puberty po menopauzu. Na rozdíl od spermatogeneze, při níž vznikají čtyři rovnocenné haploidní prekurzory spermií, oogeneze vede k jedinému zralému haploidnímu oocytu, ostatní pólová tělíska jsou menší a nefunkční.
Amitóza (přímé dělení) je z uvedených dělení nejméně rozšířená. Buňka se dělí tak, že se jádro prodlouží a rozdělí na dvě části, aniž by zanikla jaderná membrána. Následně se rozdělí i zbývající prostor mateřské buňky na dvě buňky dceřiné. Nedochází ke spiralizaci chromosomů, nelze pozorovat ani vlákna dělícího aparátu. Na rozdíl od mitózy a meiózy není v případě amitózy zaručeno rovnoměrné rozdělení genetického materiálu. Proto se u člověka vyskytuje pouze zřídka (u specializovaných buněk, např. tkáně epitelu močového měchýře) a většinou je spojena s patologickými ději (kancerogenezí). Od amitózy je nutné rozlišovat tzv. endomitózu. Jde o zvláštní typ mitózy, během níž nedochází k zániku jaderné membrány.
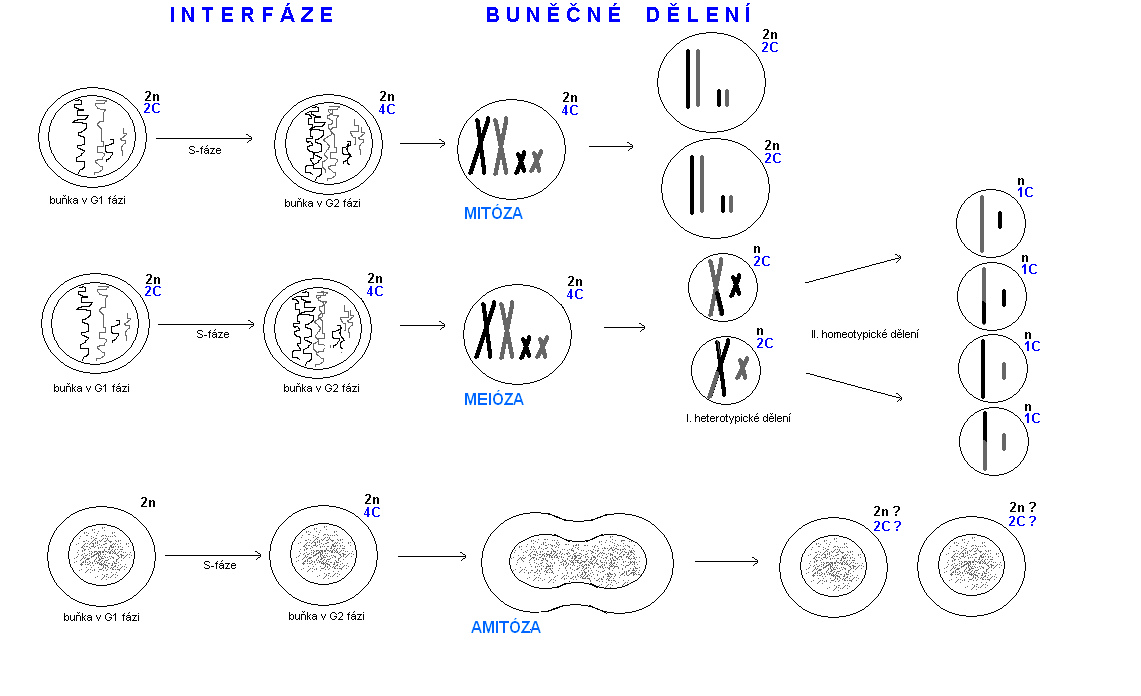
Legenda: Ačkoliv člověk obsahuje 23 párů chromosomů, pro zjednodušení a názornost jsou uvedeny pouze dva homologní páry chromosomů, které se liší velikostí. Odlišná barva (černá, resp. šedá) znázorňuje různý původ chromosomů od obou rodičů sledovaného jedince. Písmeno "n" udává ploidii (2n - diploidie, n-haploidie). Písmeno C udává absolutní počet kopií chromosomů v buňce bez ohledu na ploidii.
4.2 Rozmnožování prokaryontních buněk
4.3 Segregace a kombinace vloh
Tabulka 2 Kombinace alel krevně-skupinového systému ABO
Krevní skupina jedince |
Genotyp jedince | |
Homozygot |
Heterozygot | |
A |
IA IA |
IA i |
B |
IB IB |
IB i |
AB |
- |
IA IB |
ii |
- | |
Při důkladném prostudování tabulky 2 můžeme dojít k následujícím klíčovým zjištěním:
- Krevní skupina není „fenotypový" projev, který lze zkoumat pouhým okem, ale musí se stanovit laboratorně (např. v krevní bance).
- Jedinci s krevní skupinou A nebo B představují směs dvou různých genotypů. Jeden představuje homozygoty pro dominantní alelu (IA IA nebo IB IB) druhý heterozygoti (IA i nebo IB i), u kterého dominantní alela potlačí projev alely i.
- Jedinci s krevní skupinou AB jsou výlučně heterozygoti.
- Jedinci s krevní skupinou 0 jsou výlučně recesivní homozygoti.
Uvedený příklad krevních skupin je názornou ukázkou důsledků pohlavního rozmnožování. Jak bylo uvedeno v kapitole 4.1, pohlavní rozmnožování využívá meiózu. Při ní dochází k rozdělení - segregaci obou chromosomů homologického páru v gametách jedince. Po splynutí jeho gamety s gamety jedince opačného pohlaví vznikne nová kombinace alel u jejich potomka. Projevy dědičnosti byly zkoumány od nepaměti a lidé využívali empirické poznatky při šlechtění plemen užitkových a domácích zvířat. Vědecký základ však této oblasti poskytl až Johann Gregor Mendel, který publikoval výsledky svých pokusů s křížením hrachu (Pisum sativum L.) v roce 1865. Uplynulo však ještě více než třicet let než Hugo de Vries, Carl Correns a Erich von Tschermak znovuobjevili jeho práci na počátku 20. století.
Mendelův přínos spočívá v tom, že se pokusil empiricky pozorované projevy dědičnosti znaků definovat matematicky. Jeho zjištění, známé jako "Mendelova pravidla", jsou platné dodnes. Dříve, než si je vysvětlíme, musíme si definovat podmínky, za kterých platí. První podmínkou je, aby byly námi sledované geny lokalizovány na různých chromosomech. Kromě toho, všechny geny by měly být umístěny na somatických, nepohlavních chromosomech - autozomech. Znaky, podmíněné sledovanými alelami genů by měly být identifikovatelné a neměly by na jedince působit letálně. Pro lepší matematickou výpovědní hodnotu se dají Mendelova pravidla elegantně ověřit na živých organismech s kratší reprodukční dobou a dostatečným počtem potomstva, které vzniklo pohlavním rozmnožováním.
Dříve než se budeme věnovat Mendelovým pravidlům, je nutno uvést specifické symboly, které se při genetickém zápisu používají:
P rodičovská (parentální) generace
F1 potomstvo křížení rodičů (první filiální generace)
F2 potomstvo křížení generace F1 (druhá filiální generace, vnuci generace P)
GP gamety parentální generace (spermie, vajíčka)
GF1 gamety generace F1
B zpětné křížení potomků s jedincem identického genotypu, jaký měl jeden z rodičů v parentální generaci
♀ označení ženského pohlaví v zápise genetického křížení
♂ označení mužského pohlaví v zápise genetického křížení
x symbol křížení
R dominantní alely se obecně označují velkými písmeny
r recesivní alely se obecně označují malými písmeny
Rh-faktor u člověka je kódován geny, lokalizovanými na chromosomu č. 1 (lokace 1p34-36). Nejdůležitější z nich je gen, který kóduje antigen D. Má dvě alely, přičemž dominantní alela R je zodpovědná za tvorbu antigenu. Recesivní alela r nepodmiňuje tvorbu antigenu D v důsledku delečních změn, kterými se liší od dominantní alely R.
Představme si manželský pár. Žena je genotypově dominantní homozygot s oběma dominantními alelami RR, proto je fenotypově Rh+. Muž je recesivní homozygot rr, proto je při hematologickém vyšetření určený jako Rh-. Z obrázku 2 je patrno, že z obou kopií genů (alel) se při gametogenezi může dostat do gamety (spermie, vajíčka) pouze jedna alela, proto vajíčka ženy obsahují pouze jednu z obou dominantních alel R. Spermie muže nesou také pouze jednu z obou recesivních alel r. Bez ohledu na to, kolik bude mít uvedený pár dětí, všechny děti budou z hlediska Rh-faktoru identické. Fenotypově budou při vyšetření Rh+ a z hlediska genotypu se bude jednat o heterozygoty Rr, kteří obsahují jednu dominantní alelu R od matky a jednu recesivní alelu r od otce.
♀ ♂
P: RR x rr
GP: R r
F1: Rr
Uvedený genetický zápis je příkladem prvního Mendelova pravidla uniformity a reciprocity: Při křížení rodičů homozygotních ve sledovaném znaku (generace P) jsou všichni jejich potomci stejní - uniformní genotypově (Rr) i fenotypově (hematologické vyšetření prokáže Rh+). Zároveň je jedno, který z obou rodičů byl dominantní homozygot a který recesivní homozygot, protože se jedná o gen na somatickém chromosomu (autozomu). Všimněte si, že při genotypu ženy rr a genotypu muže RR bychom pozorovali stejné potomstvo. Jedná se tedy o vzájemnou zaměnitelnost – reciprocitu genotypů.
Poznámka: Při genetickém zápisu se vždy píše jako první dominantní alela. Alely jednotlivých genů se musí psát vždy spolu (správně: Rr, RrBB; špatně: rRBb, RbrB)!
Pokud by si dítě uvedeného páru našlo genotypově shodného partnera (heterozygota Rr, označený červeně), můžeme uvažovat o zplození další generace, druhé filiální generaci F2. V tomto případě však rodiče tvoří dva typy gamet. Polovina z nich obsahuje dominantní alelu R a druhá polovina má recesivní alelu r.
♀ ♂
F1 Rr x Rr
GF1 R ; r R ; r
½ ; ½ ½ ; ½ Pravděpodobnost vzniku gamet
F2 RR : Rr + Rr : rr
¼ ½ ¼ Pravděpodobnost vzniku potomků
1 : 2 : 1 Genotypový štěpný poměr
3 Rh+ : 1 Rh- Fenotypový štěpný poměr
Na generaci F2 můžeme sledovat druhé Mendelovo pravidlo štěpení znaků: V případě křížení heterozygotní generace F1 se v jejím potomstvu F2 znovu objeví kombinace, jaké měla rodičovská generace P. Na základě pravděpodobnosti vzniku jednotlivých gamet si můžeme vysvětlit genotypové a fenotypové štěpné poměry F2 generace potomků. Oba rodiče tvoří gamety nesoucí alelu R s pravděpodobností ½ a gamety s recesivní alelou r s pravděpodobností ½. Proto pravděpodobnost, že dítě v F2 generaci bude dominantní homozygot RR, činí ¼ (½R · ½R = ¼RR) a pravděpodobnost recesivního homozygota rr představuje rovněž ¼ (½r · ½r = ¼rr). Pravděpodobnost vzniku heterozygota Rr bude ½ ((½R · ½r) + (½R· ½r) = ½Rr). Fenotypový štěpný poměr je jiný, protože v případě heterozygotů Rr překryje dominantní alela R projev recesivní alely r, proto dítě bude vyšetřeno jako Rh+. S pravděpodobností ¾ bude generace F2 vykazovat Rh+ a pouze ¼ dětí bude Rh-.
V případě gamet obou generací je vidět princip třetího Mendelova pravidla o čistotě gamet. Gameta může z daného páru chromosomů jedince obsahovat pouze jeden chromosom.
Až dosud jsme sledovali jeden znak. Jedinci F1 generace s genotypem Rr se nazývají monohybridi, protože vznikly křížením (hybridizací) rodičovských homozygotů. Pokud sledujeme dva geny, mluvíme o dihybridismu, v případě tří znaků o trihybridismu a v případě více znaků o polyhybridismu.
Pokud bychom sledovali několik genů zároveň, bude platit čtvrté pravidlo o nezávislé kombinaci genů. Alely jednoho sledovaného genu se dědí a kombinují nezávisle na alel druhého sledovaného genu. Názorným příkladem je sledování krevně-skupinového systému ABO (gen se nachází na chromosomu č. 9, lokace 9q34.1-34.2) a Rh-faktoru: Pro názornost jsou oba geny označeny jinou barvou.
Křížení parentální generace: ♀ ♂
P: RRIAIA x rrii
Rh+, krevní skupina A Rh-, krevní skupina O
Gamety rodičovské generace:
GP: RIA ri
F1: RrIAi
Rh+, krevní skupina A
Křížení generace F1:
F1: RrIAi x RrIAi
GF1: RIA ; Ri ; rIA; ri RIA ; Ri ; rIA; ri
Tabulka 3 Kombinační tabulka vzniku generace F2 z gamet generace F1
Gamety generace F1 |
Žena | ||||
RIA |
Ri |
rIA |
ri | ||
Muž |
RIA |
RRIAIA Rh+, krevní skupina A |
RRIAi Rh+, krevní skupina A |
RrIAIA Rh+, krevní skupina A |
RrIAi Rh+, krevní skupina A |
Ri |
RRIAi Rh+, krevní skupina A |
RRii Rh+, krevní skupina O |
RrIAi Rh+, krevní skupina A |
Rrii Rh+, krevní skupina O | |
rIA |
RrIAIA Rh+, krevní skupina A |
RrIAi Rh+, krevní skupina A |
rrIAIA Rh-, krevní skupina A |
rrIAi Rh-, krevní skupina A | |
ri |
RrIAi Rh+, krevní skupina A |
Rrii Rh+, krevní skupina O |
rrIAi Rh-, krevní skupina A |
rrii Rh-, krevní skupina O | |
Legenda: Kombinací gamet generace F1 (modré pozadí) vznikne generace F2 (bílé pozadí). U každého genotypu generace F2 je uveden i fenotyp.
Důležitá poznámka: Všechny čtyři typy gamet (RIA; Ri; rIA; ri) se u obou pohlaví F2 generace tvoří se stejnou pravděpodobností 1: 1: 1: 1 (čili ¼ + ¼ + ¼ + ¼ = 1).
Genotypový štěpný poměr: 1 : 2 : 1 : 2 : 4 : 2 : 1 :2 :1
V tabulce jsou zastoupeny genotypové kategorie:
1x RRIAIA
2x RRIAi
1x RRii
2x RrIAIA
4x RrIAi
2x Rrii
1x rrIAIA
2x rrIAi
1x rrii
Fenotypový štěpný poměr: 9 : 3: 3: 1
9x Rh+, krevní skupina A
3x Rh+, krevní skupina O
3x Rh-, krevní skupina A
1x Rh-, krevní skupina O
Poznámka: Součet 1 + 2 + 1 + 2 + 4 + 2 + 1 + 2 + 1 = 16, což představuje kombinaci 4 typů gamet obou rodičů (kombinační tabulka 4 x 4 = 16). Součet fenotypových tříd 9 + 3 + 3 + 1 = 16.
Pravidlo o nezávislé kombinaci alel obou sledovaných genů si můžeme ověřit přímo v kombinační tabulce 3. Pokud budeme sledovat pouze gen pro Rh-faktor, v tabulce zjistíme přítomnost 4 dominantních homozygotů RR, 8 heterozygotů Rr a 4 recesivních homozygotů rr. Po vydělení čtyřmi dostaneme základní genotypový štěpný poměr platný pro monohybridismus: 1: 2: 1 (tabulka 3). Obdobně to platí i pro systém ABO.
Na závěr této kapitoly je třeba uvést několik zajímavých poznámek k tabulce 2.
- Krevně-skupinový systém ABO nám ukazuje, že i když gen má v populaci více než dvě alely, u jedince mohou být přítomny pouze dvě z nich. Např. v uvedeném příkladu matky s genotypem IAIA a otce s genotypem ii už není místo pro třetí alelu IB. Naopak, jedinec s genotypem IBIB už nemůže mít žádnou ze zbývajících alel IA nebo i.
- Z tabulky 2 vyplývá, že rodič s krevní skupinou AB (genotyp IAIB) nikdy nemůže mít dítě s krevní skupinou 0 (genotyp ii). Rodič s krevní skupinou 0 může mít dítě se shodnou krevní skupinou 0 s partnerem s krevní skupinou 0, nebo s partnerem s krevní skupinou A (v heterozygotním stavu IAi), případně B (v heterozygotním stavu IBi).
- V případě rodičů s genotypy IAi x IBi jsou u jejich dětí teoreticky možné všechny čtyři krevní skupiny (genotypy: IAIB; IAi; ii; IBi).
4.4 Vazba genů
Představme si situaci, že se dva hypotetické geny M a R nacházejí na stejném chromosomu. V takovém případě lze jedince heterozygotní pro oba znaky zapsat dvěma způsoby:
M R nebo M r
m r m R
Vidíme, že existují dvě možnosti. Vlevo je případ, kdy jsou obě dominantní alely M a R na jednom chromosomu, přičemž na druhém chromosomu jsou pouze recesivní alely m, r. Takové uspořádání alel se nazývá "fáze cis". Vpravo je schéma jedince, který má na jednom chromosomu homologického páru dominantní alelu M a recesivní r a na druhém chromosomu obráceně. Na rozdíl od jedince vlevo má na jednom chromosomu různé typy alel - i dominantní i recesivní. Uvedené uspořádání alel se nazývá "fáze trans".
Bylo by obrovskou chybou, pokud bychom toto uspořádání zapsali "mendelistickým" zápisem MmRr. Je sice pravda, že tento zápis formálně bere v úvahu dominantní i recesivní alely obou genů, ale není z něj jasná klíčová věc - že geny jsou umístěny na stejném chromosomu, tedy ve vazbě. Dvojitá čára mezi geny znázorňuje dva homologní chromosomy jediného páru, na kterém jsou umístěny. Pokud by neexistovala rekombinace, oba jedinci by produkovali v důsledku vazby genů ne čtyři typy gamet, ale pouze dva. Pokud by však mezi geny vznikl crossing-over, vznikly by další dva typy gamet. Protože proces rekombinace je náhodný, počet vzniklých gamet s "rekombinovanými" geny M a R bude vždy nižší ve srovnání s "rodičovskými" kombinacemi genů M a R v gametách, které vznikly jednoduchou segregací chromosomů: Všimněte si, že fáze cis a trans se liší v typu "rodičovských" a "rekombinovanými" gamet - jsou recipročně obrácené.
Jedinci:
M R nebo M r
m r m R
fáze cis fáze trans
Gamety:
M R M r rodičovské gamety
m r m R rodičovské gamety
M r M R rekombinované gamety
m R m r rekombinované gamety
Jak zjistíme, zda má jedinec uspořádané geny ve fázi cis nebo trans? Řešení nám naznačuje poznatek, že na rozdíl od volné kombinovatelnosti genů nebude počet jednotlivých typů gamet shodný, ale nerekombinované rodičovské sestavy budou v gametách vždy početnější. Tuto skutečnost dokáže odkrýt tzv. zpětné křížení (B) testovaného jedince s dvojitým recesivním homozygotem.
B M R X m r M r X m r
m r m r m R m r
fáze cis fáze trans
GB M R; m r; M r; m R m r M R; m r; M r; m R m r
B1
M R M r
m r m r
m r m R
m r m r
M r M R
m r m r
m R m r
m r m r
Vidíme, že recesivní homozygot je při tomto křížení důležitý proto, že tvoří stejné gamety s recesivními alelami m r i v případě, že mezi geny vznikl crossing-over. Kromě toho, v případě křížení heterozygota s recesivním homozygotem jsou genotypové počty rozlišitelné i fenotypově. Jinými slovy, v případě zpětného křížení jsou genotypové i fenotypové štěpné poměry shodné.
Splynutím rodičovských gamet generace B vzniknou jedinci v generaci potomků B1. Černou barvou jsou označeni jedinci, kteří od heterozygotního rodiče zdědili původní, tedy nerekombinované rodičovské kombinace alel. Modrou barvou jsou označeni potomci, kteří vznikli z gamet, které obsahovaly rekombinovanou sestavu genů. Těchto "rekombinovaných" jedinců je vždy méně než jedinců s původní rodičovskou sestavou alel.
Míra rekombinace se dá využít k určení vzdálenosti mezi geny, které jsou ve vazbě. Pokud bychom udávali vzdálenosti mezi nimi v absolutních počtech párů bází, čísla by dosahovaly obrovské numerické hodnoty. Proto je výhodnější určit frekvenci rekombinace. Obecně platí, že čím jsou geny k sobě blíž, tím menší je pravděpodobnost vzniku crossing-overu. Pokud budeme sledovat dva geny v potomstvu zpětného křížení, můžeme určit jejich vzájemnou vzdálenost, která je udávána v centimorganech (cM) a představuje procento jedinců potomstva, kteří nesou rekombinované sestavy sledovaných genů. Nazývá se Morganovo číslo p a je určeno následujícím vztahem:
p(%; cM) = (počet rekombinantů / počet všech jedinců) · 100
Kromě toho můžeme určit, s jakou frekvencí se častěji tvoří rodičovské nerekombinované sestavy alel sledovaných genů v porovnání s uspořádáním, které vzniklo díky crossing-overu. Tato frekvence se nazývá Batesonovo číslo c a platí pro ni vztah:
c(x) = (počet rodičovských kombinací/počet rekombinací)
Mezi oběma čísly platí přepočítací vztahy:
p = 100/(c + 1) c = (100 – p)/p
Příklad 1: Předpokládejte, že u laboratorního potkana jsou geny N a E ve vazbě. Určete vzdálenost mezi nimi. Máte k dispozici počty jednotlivých genotypových i fenotypových kategorií v potomstvu.
B N E X n e
n e n e
N E 40
n e
n e 30
n e
N e 16
n e
n E 14
n e
Řešení: V generaci potomstva křížení heterozygota jsme pozorovali 70 jedinců s nerekombinovanými rodičovskými sestavami a 30 jedinců, kteří vznikli rekombinací genů N a E. Po dosazení do vztahu pro výpočet Morganová čísla p dostáváme:
p = (30/100) · 100 c = 70/30
p = 30 % (cM) c = 2,33x
Vzdálenost mezi geny činí 30 cM, procento jedinců potomstva, kteří nesou ve svém genotypu rekombinované sestavu alel sledovaných genů, představuje 30 %. Rodičovské kombinace se v gametách heterozygotů vyskytují 2,33 krát častěji než rekombinovanými sestavami. V generaci B1 pozorujeme, že vyšší počet jedinců se genotypově i fenotypově podobá oběma rodičům. Potomků, kteří mají pouze jednu dominantní alelu v genotypu, je méně, protože všichni vznikli výhradně díky crossing-overu. Proto se fáze cis nazývá také "coupling" (z angl. coupling - spojení, vazba).
Příklad 2: Předpokládejte, že u laboratorního potkana jsou geny M a R ve vazbě. Určete vzdálenost mezi nimi. Máte k dispozici počty jednotlivých genotypových a fenotypových kategorií v potomstvu.
B M r X m r
m R m r
B1 Genotyp: Zjištěné počty jedinců:
M r 40
m r
m R 30
m r
M R 16
m r
m r 14
m r
Řešení: V generaci potomstva křížení heterozygota jsme pozorovali 80 jedinců s nerekombinovanými rodičovskými sestavami a 20 jedinců, kteří vznikli rekombinací genů M a N. Po dosazení do vztahu pro výpočet Morganová čísla p dostáváme:
p = (20/100) · 100 c = 80/20
p = 20 % (cM) c = 4,0x
Vzdálenost mezi geny činí 20 cM, procento jedinců potomstva, kteří nesou ve svém genotypu rekombinovanou sestavu alel sledovaných genů, představuje 20 %. Rodičovské kombinace se v gametách heterozygotů vyskytují 4 krát častěji než rekombinované sestavy. Jedinců v B1 generaci, kteří se genotypově a fenotypově podobají oběma rodičům, je méně, než bychom očekávali. Je to typické pro heterozygotní rodiče ve fázi trans, která se proto nazývá také "repulsion" (z angl. repulsion – odpuzování, averze).
4.5 Genetická informace a pohlaví
Uvedený poměr však u člověka není neměnný. Primární poměr pohlaví (poměr pohlaví zygot těsně po splynutí gamet) je posunut výrazně ve prospěch heterogametického pohlaví. Sekundární poměr pohlaví se stanoví při narození dětí a činí 100 děvčat: 105 chlapců. Terciární poměr pohlaví je zjišťován v pozdějších stadiích postnatálního vývoje. U gerontů činí přibližně 100 žen: 50 mužům.
I když jsou gonozomy při chromosomovém určení pohlaví důležité, nejsou jedinými faktory, které ho podmiňují. Důležitou roli hrají i tzv. feminní a maskulinní faktory. Jsou to genové komplexy, které se nacházejí jak na monozomech, tak i na autozomech a spolupodílejí se na vývoji pohlaví. Mluvíme o genotypovém určení pohlaví.
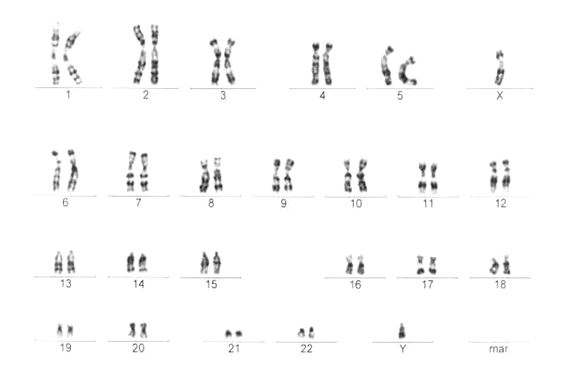
Obrázek 4 - Karyotyp muže
Morfologické rozdíly mezi gonozomy X a Y musíme brát v úvahu v případě genů, které se na nich nacházejí. Chromosom X (a v menším rozsahu i chromosom Y) obsahují oblast, ve které nejsou shodné - homologní. V případě, že se geny nacházejí v nehomologní oblasti gonozomu, mluvíme o znacích úplně vázaných na pohlaví. Příkladem je dědičnost hemofilie. Je to porucha srážlivosti krve, podmíněná mutací genu pro koagulační faktor, který se nachází na nehomologním úseku chromosomu X. Tento úsek nemá shodný protějšek na chromosomu Y. Proto se u muže gen pro koagulační faktor nachází pouze v jediné kopii na chromosomu X, zatímco u ženy je přítomen ve dvou kopiích na obou chromosomech X. Mohou nastat následující kombinace:
Genotyp Fenotyp
XHXH zdravá žena s oběma zdravými alelami
XHXh zdravá žena s jednou mutantní alelou (přenašečka, konduktorka)
XhXh žena s oběma mutantními alelami (nemocná na hemofilii)
XHY zdravý muž s jedinou zdravou alelou na chromosomu X
XhY nemocný muž s jedinou mutantní alelou na chromosomu X
Z uvedených zápisů genotypů vyplývají důležité závěry:
- Všimněte si prosím způsobu genetického zápisu. Úplnou vazbu genu na pohlaví zaznamenáváme tak, že jako hlavní znak zapíšeme symbol příslušného gonozomu (X nebo Y) a jako jeho pravý dolní index píšeme vlastní symbol konkrétní alely sledovaného genu. Vazba na pohlaví tedy znamená, že ve skutečnosti jsme nuceni sledovat znaky dva - vlastní gen a chromosomovou determinaci pohlaví jedince.
- Muž nikdy nemůže být heterozygotní pro gen, který je zcela vázán na nehomologní úsek chromosomu X, protože může mít v genotypu pouze jediný chromosom X. Z toho vyplývá, že pro sledovaný gen, který je zcela vázán na nehomologní úsek X, obsahuje pouze jednu jedinou alelu. Tento zvláštní případ u muže se nazývá hemizygotní stav.
Pro geny umístěny na nehomologních úsecích chromosomu X je typická tzv. dědičnost křížem. V případě, že žena je pro sledovaný znak recesivní homozygot, v další generaci zdědí její fenotyp opačné pohlaví - tedy její syn. Příkladem může být dědičnost daltonizmu. Je to porucha vnímání zvláště červené a zelené oblasti barevného spektra. Gen je lokalizován na nehomologním úseku chromosomu X. Pokud je matka recesivně homozygotní pro sledovaný znak (s poruchou barvocitu) a muž obsahuje na svém jediném chromosomu X zdravou alelu, jejich děti nebudou fenotypově uniformní. Dcery budou zdravé (ovšem heterozygotní přenašečky) a synové nebudou rozeznávat červenou a zelenou barvu.
Poznámka: Všimněte si, že označování pohlaví symboly ♀, ♂ je zde zcela zbytečné.
Dědičnost křížem – daltonismus:
P XdXd x XDY
„barvoslepá“ „zdravý“
GP Xd XD ; Y
F1 XDXd XdY
„zdravá“ „barvoslepý“
Pro geny umístěny na nehomologním úseku chromosomu Y je typická přímá dědičnost, nazývaná i dědičnost holandrická. Znak přechází z jednoho mužského potomka na dalšího a vždy se fenotypově projeví. Příkladem je gen, který podmiňuje hypertrichosis auriculae. Je to nadměrné ochlupení horní a zadní části ušního boltce. Vyskytuje se výlučně u mužů.
Dědičnost přímá – holandrická:
P XX x XYH
„hypertrichosis auriculae“
GP X X ; YH
F1 XX XYH
„hypertrichosis auriculae“
V některých učebnicích genetiky se uvádí, že dědičnost genů, které jsou lokalizovány na homologních úsecích chromozomů X a Y se řídí stejnými pravidly, jako dědičnost genů lokalizovaných na autozomech. Důrazně upozorňujeme, že to sice pravda je, ovšem není úplná. Názorně nám to vysvětlí příklad hypotetických genů A a B u laboratorního potkana, které jsou lokalizovány v homologním úseku gonozomů. V tomto případě říkáme o znacích neúplně vázaných na pohlaví.
Neúplná vazba genu na pohlaví - dvě možnosti dominantního a recesivního homozygota vzhledem k pohlaví:
P XAXA x XaYa anebo XaXa x XAYA
dominantní recesivní recesivní dominantní
homozygot homozygot homozygot homozygot
GP XA Xa; Ya Xa XA; YA
F1 XAXa x XAYa XAXa x XaYA
GF1 XA; Xa XA; Ya XA; Xa Xa; YA
F2 XAXA ; XAXa ; XAYa ; XaYa XAXa ; XaXa ; XAYA ; XaYA
Můžeme vidět, že generace F1 je sice v obou případech uniformní a platí i reciprocita parentální generace, ale v generaci F2 je recesivní homozygot stejného pohlaví, u jakého byly obě recesivní vlohy v parentální generaci.
Další důležitou kategorií jsou znaky pohlavně ovládané. Geny, kterými jsou kódované, jsou umístěny na autozomech, ale jejich fenotypový projev je výrazně odlišný v závislosti na pohlaví organismu, ve kterém se nacházejí. Týká se to především určení pohlavního dimorfizmu, jak je tomu v případě sekundárních pohlavních znaků. Jejich genetický základ na autozomech je shodný, ale účinkem hormonální regulační kaskády se fenotypově projeví pouze u jednoho z obou pohlaví.
U člověka (u savců obecně) zůstává u homogametického pohlaví jeden ze dvou chromosomů X spiralizován i v průběhu interfáze. Spiralizace víceméně podmiňuje inaktivaci chromosomu, resp. většiny genů, které jsou na něm umístěny. Inaktivní chromosom je možno pozorovat cytogenetickými mikroskopickými technikami v jádru buňky i během interfáze jako tzv. Barrovo tělísko. Tento jev se nazývá lyonizace.
Na závěr této kapitoly musíme upozornit na zajímavou skutečnost. Způsob chromosomového určení pohlaví u člověka není v přírodě jediný a v průběhu evoluce se vytvořily až tři typy:
- Typ Drosophila je charakteristický pro člověka, savce, většinu druhů rostlin, hmyzu a pro některé druhy ryb a plazů. Má genotypové určení pohlaví XX, XY. Název typu je odvozen od octomilky (Drosophila melanogaster).
- Typ Abraxas je charakteristický tím, že heterogametickým pohlavím jsou samice (XY nebo ZW), zatímco homogametickým pohlavím jsou samci (XX nebo ZZ). Tento typ se vyskytuje především u ptáků a jeho název je odvozen od píďalky angreštové (Abraxas grossulariata).
- Typ Habrobracon je od obou předešlých typů odlišný v tom, že samičí pohlaví je diploidní (2n), zatímco samci mají haploidní genotyp (n). Vyskytuje se u některých druhů společenského hmyzu (např. u včel).
Z hlediska genového určení pohlaví je důležitá lokalizace maskulinních a feminních faktorů. Také v tomto případě rozeznáváme tři typy.
- Typ Drosophila má maskulinní faktory umístěny na autozomech a feminní faktory na gonozomu X.
- Typ Lymantria má feminní faktory lokalizovány na heterochromosomu Y nebo W, přičemž jejich vliv u heterogametického pohlaví převládá nad maskulinními faktory, které jsou lokalizovány na chromosomech X nebo
- Typ Habrobracon má oba typy faktorů - maskulinní i feminní lokalizovány na chromosomu X. O jejich projevu rozhoduje ploidita organismu.
5 Mutace a poškození genetické informace
5.1 Klasifikace mutací
A/ Standardní stav: Uvažujme o chromosomu, na kterém sledujeme 8 genů (M, N, O, P, R, S, T, U), jejich postavení vůči centromeře C a oběma koncům chromosomu - telomerám T. Organismus obsahuje za normálních okolností dva chromosomy, které tvoří homologický chromosomový pár s uspořádáním:
Všechny uvedené chromozomální aberace představují zásadní problém, protože dochází k přestavbě genetického materiálu, čímž se zvyšuje riziko postižení potomstva. Chromosomy s aberacemi nejsou schopny dokonalého homologního párování v průběhu mitózy a meiózy, protože se liší od standardního uspořádání chromosomu. V případě chromosomů bez centromery (acentrických) dochází v průběhu buněčného dělení k nevratné ztrátě genetické informace, protože právě centromera zaručuje správný přenos obou kopií chromosomů s pomocí dělícího aparátu do dceřiných buněk.
Musíme si uvědomit fakt, že v buňce není jediná mitochondrie ale stovky až tisíce, přičemž mutace nemusí být stejná u všech. V tomto případě mluvíme o heteroplazmii. Homoplazmie označuje stav, kdy jsou v buňce přítomny mitochondrie se shodnou genetickou informací.
5.2 Mutagenní faktory
5.3 Reparační mechanismy
6 Populační genetika
6.3 Testovací otázky
6.1 Kvalitativní znaky v populaci
Četnosti:
|
A
p |
a
q |
A
p
|
AA
p2
D |
Aa
pq
H |
a
q
|
Aa
pq
H |
aa
q2
R |
V jmenovateli je dvojnásobek počtu všech jedinců populace, protože každý jedinec obsahuje dvě alely pro sledován gen (viz. diploidní stav, kapitola 4).
6.2 Kvantitativní znaky v populaci

Poznámka: Příklad PTC jsme nevybrali náhodně. Vzhledem k hořké chuti některých přírodních látek i farmak může představovat míra rozeznání této chutě v závislosti od koncentrace látky u pacientů důležitý faktor při terapii.
P = G + E (6)
Část variability sledovaného znaku, která je podmíněna geneticky, vyjadřuje tzv. dědivost (heritabilita) h2 podle vztahu:
h2 = σ2G/ σ2P (7)
7 Aplikace genetiky ve zdravotnictví
7.1 Genealogické vyšetření
Poznámka: V používání symboliky existují ve světě drobné rozdíly, ty však nejsou natolik závažné, aby bránily pochopit znázorněný rodokmen, proto se jim blíže nebudeme věnovat.
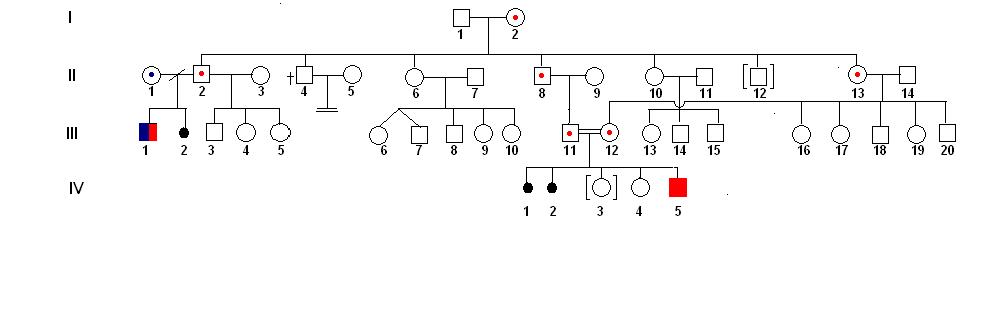
Cílem této kapitoly je vysvětlit povahu dědění jednotlivých znaků. Proto si vytvoříme hypotetický rodokmen (obrázek 6). Na tomto rodokmenu si ukážeme typický projev autozomálních dominantních alel, autozomálních recesivních alel, dominantních alel vázaných v nehomologním úseku chromosomu X, recesivních alel vázaných v nehomologním úseku chromosomu X a matroklinní dědičnosti, kterou musíme brát v úvahu v případě mitochondriálních genů.
Poznámka: Je jasné, že v jediném rodokmenu se nemohou vyskytovat všechny uvedené typy, které jsou vyznačeny na obrázku 6. Jsme však toho názoru, že jediná hypotetická schéma rodokmenu může napomoci rychlejšímu pochopení podstaty jednotlivých typů dědičnosti bez nutnosti nanovo pochopit příbuzenské vztahy, která by byla nutná v případě odlišných rodokmenů.
Jednotlivé generace se v rodokmenu označují římskými čísly. Jedinci se v rámci téže generace označují arabskými čísly podle pořadí narození. Např. v hypotetickém rodokmenu (obrázek 6A) první generaci tvoří manželský pár I1 a I2. Měli šest vlastních biologických dětí (II2, II4, II6, II8, II10 a II13) a jednoho adoptivního syna (II12). Jejich vnuk (III3) pochází z prvního manželství nejstaršího syna (II2). Po rozvodu se nejstarší syn podruhé oženil a měl tři děti, prvního syna (III3) a dvě dcery (III4 a III5). Kluk III1 je polorodý sourozenec dětí III3, III4 a III5 (stejný otec, různé matky). V rodině je příbuzenský sňatek mezi bratrancem a sestřenicí prvního stupně (III11 a III12), přičemž otec muže (II8) je starším bratrem matky jeho manželky, která je současně jeho sestřenicí (II13). Uvedený příbuzenský pár měl dva neúspěšné pokusy o dítě, které skončily spontánním abortem (IV1, IV2). Následně si pár adoptoval děvčátko (IV3). Po využití technik asistované reprodukce se jim narodily dvě děti, starší holčička (IV4) a nejmladší chlapeček (IV5).

Autozomálně recesivní vlohy mají tyto specifika (obrázek 6C):
Po přehledem vysvětlení povahy jednotlivých typů dědičnosti při sledování rodokmenu si musíme objasnit důsledky příbuzenského křížení. Pokud se vrátíme k vysvětlení autozomálně recesivního typu dědičnosti, víme, že převážná část mutantních alel je v heterozygotním stavu a proto se jimi podmíněné onemocnění neprojevuje. Porucha vzniká pouze v případě, pokud se u jedince setkají dvě mutantní alely od obou heterozygotních rodičů. Obě recesivní alely v tomto případě mohou, ale také nemusí být identické původem (IBD). Např. v případě fenylketonurie (kapitola 5.1 Klasifikace mutace) známe několik různých mutantních alel. Je to logické, protože mutace může zasáhnout různá místa příslušného genu, přičemž výsledek je stejný - znefunkčnění enzymu fenylalaninhydroxylázy.
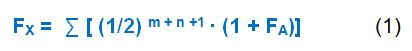

Poznámka: Při výpočtu inbredního koeficientu probanda se za nulovou generaci pokládá generace jeho rodičů, jako první je generace jeho prarodičů, druhá bude generace praprarodičů, atd. Pro uvedený příklad červeně označeného probanda na obrázku 7 činí inbrední koeficient Fx = ∑ [ (1/2) 2 + 2 +1 . (1 + 0)] = 0,03125. Výsledek platí ovšem pouze za předpokladu, že hodnota inbredního koeficientu společného předka FA=0 (tj. nepochází z příbuzenského svazku).
7.2 Cytologická analýza
Základním zpracováním chromosomů je G-proužkování (využívá aplikaci trypsinu a Giemsova barviva. Kromě toho známe R-proužkování, při kterém je obrazec vzniklých proužků reverzní (jakoby fotografický negativ) vůči G-proužkování. Při C-proužkování jsou velmi dobře pozorovatelné centromery chromozomů. Q-proužkování využívá chiakridin a při NOR-proužkování se chromosomy vizualizují s pomocí AgNO3.
Techniky proužkovaný chromozomů HRT (z angl. high resolution techniques) využívají zastavení dělení buněk před metafází (tzv. prometafáze, na rozhraní profáze a metafáze). Proto jsou chromosomy méně spiralizované, jsou delší, jemnější a lze pozorovat více pruhů (cca 800).
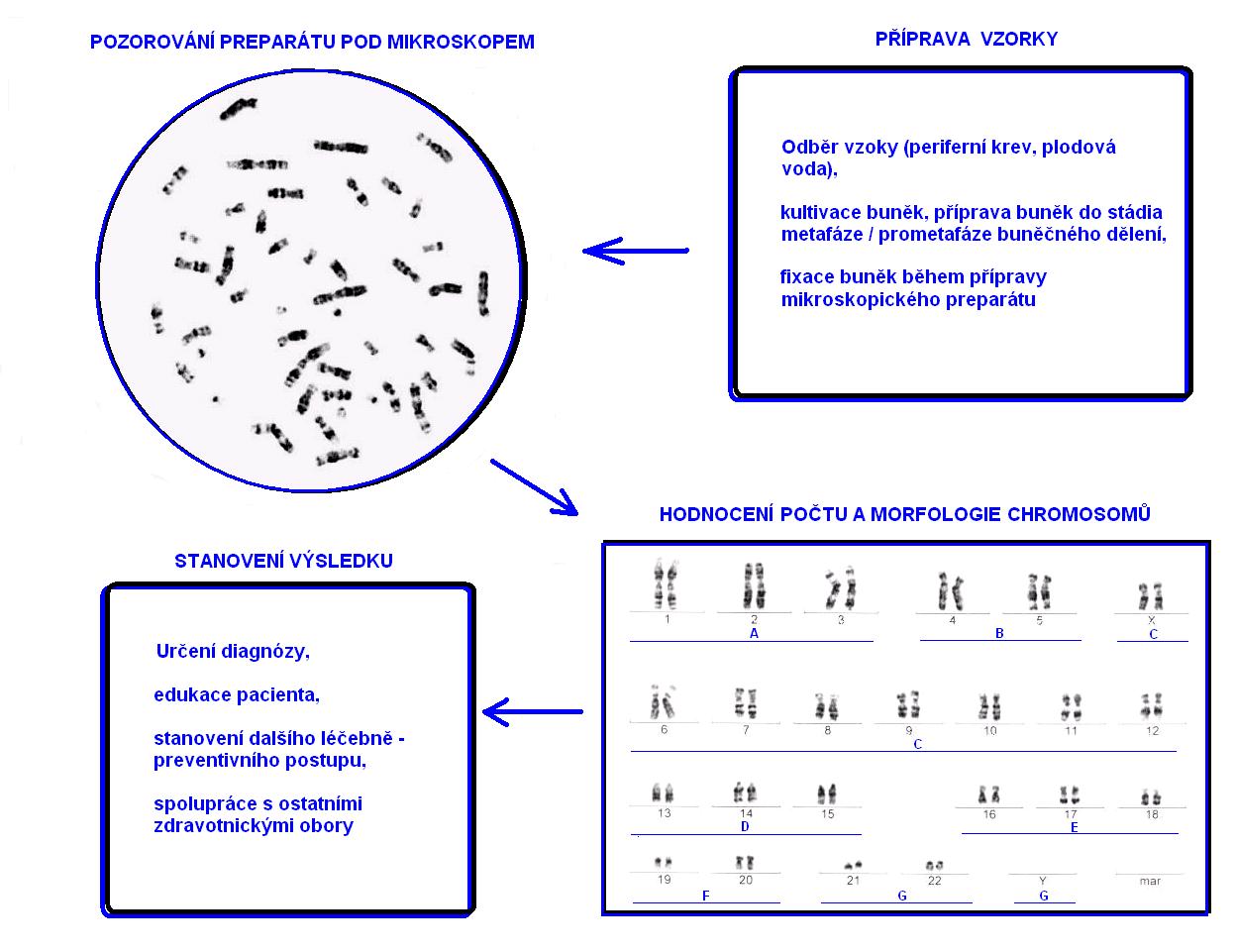
Charakteristika
|
Skupina chromosomů | ||||||
A |
B |
C |
D |
E |
F |
G | |
Autozomy |
1 - 3 |
4 – 5 |
6 – 12 |
13 – 15 |
16 – 18 |
19 – 20 |
21- 22 |
Gonozomy |
- |
- |
X |
- |
- |
- |
Y |
Počet u muže |
6 |
4 |
15 |
6 |
6 |
4 |
5 |
Počet u ženy |
6 |
4 |
16 |
6 |
6 |
4 |
4 |
Skupina chromosomů |
Charakteristika |
A |
Velké metacentrické chromosomy (centromery víceméně uprostřed chromosomu, obě raménka chromosomu stejně dlouhé) |
B |
Velké submetacentrické chromosomy (centromery posunuta mimo střed chromosomu, vznik kratšího raménka „p“ a delšího raménka „q“) |
C |
Středně velké metacentrické chromosomy |
D |
Středně velké akrocentrické chromosomy (centromery poblíž konců chromosomů-telomer)
|
E |
Malé submetacentrické chromosomy |
F |
Malé metacentrické chromosomy |
G |
Malé akrocentrické chromosomy |
7.3 Biochemická genetika
Garrod si jako první všiml, že uvedený soubor symptomů je přenášen v rodokmenu postižených probandů. Jinými slovy, jedná se o genetickou vlohu. Vzhledem k tomu, že Mendelovy zákony byly v té době právě "znovuobjeveny" a širší odborné veřejnosti stále v podstatě neznámé, podobně jako biochemická podstata metabolických dějů člověka, také Garrodův objev zůstal na několik desetiletí nedoceněný.
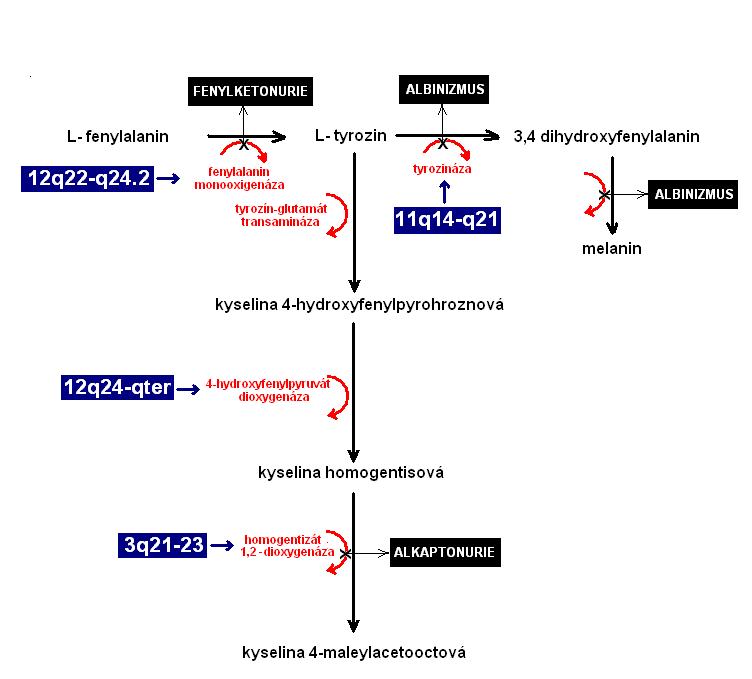
Dnes víme, že alkaptonurie je autozomálně recesivní dědičné onemocnění, jehož podstatou je ztráta aktivity enzymu homogentizát 1,2 dioxygenázy. Její gen se nachází u člověka na delším raménku chromozomu č. 3, lokace 3q21-23. Defektní enzym mohou způsobit mutace na více místech nukleotidové sekvence genu. Dnes jich je známých několik desítek. Většina mutantních (recesivních) alel je skryta u heterozygotů, kteří jsou metabolicky zdraví, protože na druhém chromozomu mají standardní "zdravou" alelu. Pokud se u jedince setkají dvě mutantní alely (které nemusí být identické), jedinec je postižený alkaptonurií. V naší populaci je frekvence tohoto onemocnění přibližně 1: 20000 novorozenců.
Příklad alkaptonurie jsme neuvedli náhodou a chceme jím zdůraznit provázanost biochemických metabolických drah člověka. Tato dědičná choroba je důsledkem selhání jednoho z enzymů téže dráhy, kde v jednom z kroků vzniká mutací i fenylketonurie, kterou jsme se podrobně zabývali v kapitole 5.1 Klasifikace mutací (Příklad 1).
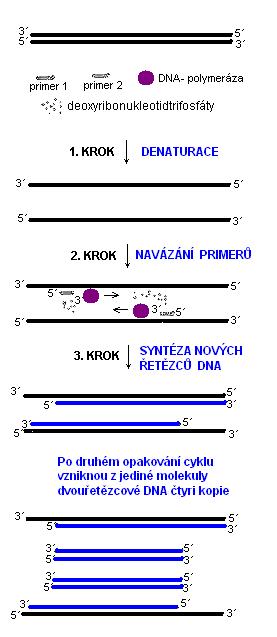
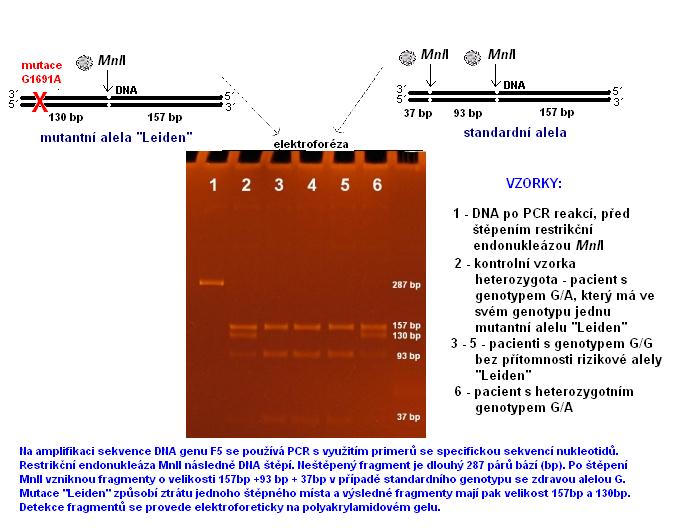
7.4 Molekulárně genetické metody
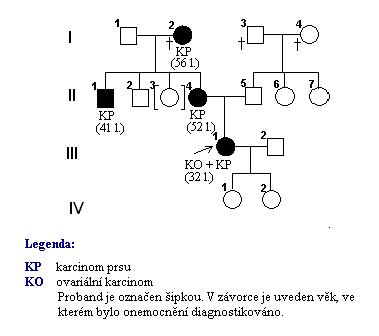
Genealogická analýza byla v tomto případě důležitá nejen z hlediska managementu léčebně preventivního procesu u pacientky s kombinovaným karcinomem prsu a ovarií. Jak můžeme vidět na obrázku 12, 32- letá pacientka byla vdaná a měla dvě děti, obě dcerky. Vzhledem k familiárnímu výskytu onemocnění karcinomu prsu v rodině je vyšetření těchto dívek velmi důležité pro stanovení prognózy možného vzniku onemocnění.
1 / Není přítomen žádný HPV. Vysvětlení může být dvojí. Nejpravděpodobnější je, že vyšetřená žena ještě neměla pohlavní styk, nebo při pohlavním styku nebyla infikovaná žádnou variantou HPV. V tomto případě však musíme brát v úvahu i možnost tzv. falešné negativity, způsobené např. nevhodným nebo nesprávným odběrem vzorku v ambulanci gynekologa, který negativně ovlivní i výsledek laboratorního testování.
Interní řízení kvality (angl. Internal quality control), často nesprávně překládané jako interní kontrola kvality) spočívá v používání vzorků se známým genotypem, které jsou vyšetřovány v sérii spolu s neznámými vzorky pacientů. Pokud je výsledkem analýzy jiný genotyp kontrolního vzorku, než jaký byl ověřen, analýza se musí zopakovat. Kromě této tzv. pozitivní kontroly se používá i negativní kontrola. Její principem je provedení celé molekulárně-genetické analýzy na vzorku, který je zcela shodný s ostatními s jedinou výjimkou - neobsahuje DNA. Pokud přesto, že ve vzorku není DNA, získáme jakýkoliv výsledek, je zřejmé, že došlo ke kontaminaci vzorku a molekulárně-genetické vyšetření je nutno zopakovat.
Externí řízení kvality (angl. external quality control) spočívá ve vyšetření vzorku, jehož genotyp je laboratoři neznámý, avšak poskytovateli kontroly známý je. Cílem je ověřit, zda laboratoř opravdu stanoví takový genotyp, jaký stanovit má.
Genetická mozaika je stav, kdy se u pacienta nacházejí buňky a tkáně s různým genetickým materiálem. Většinou je to v důsledku somatické mutace během ranní embryogeneze. Platí, že čím dříve se v průběhu vývoje jedince mutace vyskytne, tím větší počet somatických buněk ji bude obsahovat. Přirozený výskyt mozaicizmu (tj. bez mutační změny) je u člověka velmi vzácný a byl popsán v případech anomálií vzájemného ovlivnění plodů (např. dvojčat) v průběhu těhotenství.
Poznámka: Je samozřejmě možné použít v případě nouze i jiný odběrový materiál, ale bylo by vhodné to předem konzultovat. Naopak, pracovníci laboratoří by měli mít přehled o odběrových systémech na trhu, aby věděli v takovém případě erudovaně poradit.
8 Budoucnost využití genetiky ve zdravotnictví
8.4 Testovací otázky
8.1 Nanotechnologie
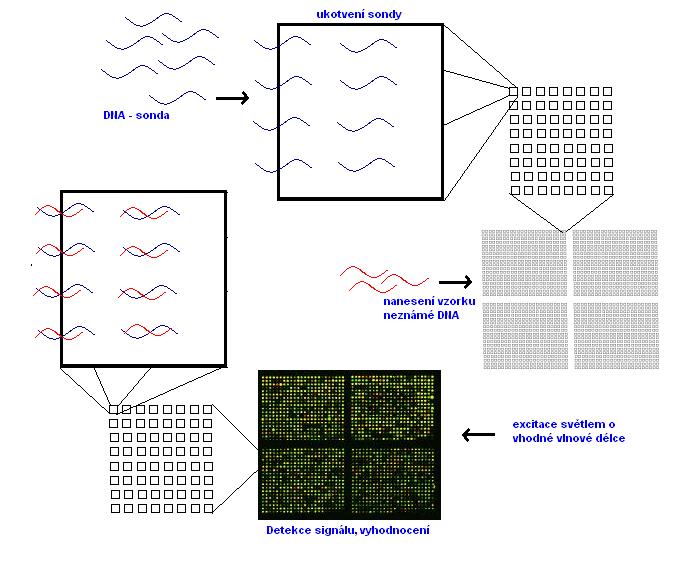

8.2 Molekulární medicína
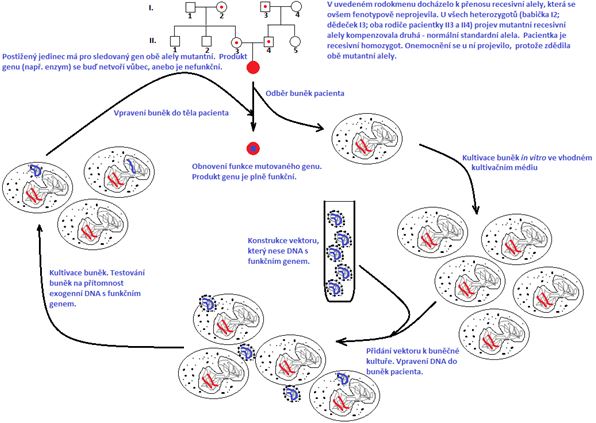
8.3 Etické problémy
Princip |
Požadavky |
Beneficience |
Veškeré diagnostické a terapeutické postupy musí sloužit k prospěchu pacienta. |
Non-maleficience |
V průběhu diagnostiky i terapie pacienta se zdravotničtí pracovníci musí zdržet jednání, které je spojeno s rizikem poškození pacienta (např. prodlužování života pacienta v terminálním stadiu nemoci za každou cenu bez ohledu na jeho utrpění). |
Spravedlnosti |
Diagnostika a terapie je u všech pacientů vykonávána spravedlivě, bez diskriminace. |
Autonomie lidské bytosti |
Zdravotník musí respektovat jedinečnost pacienta jako lidské bytosti, která má právo rozhodovat o svém zdraví. |
Solidarity |
Systém zdravotnictví musí poskytnout přiměřenou diagnostiku a léčbu všem, kteří to potřebují. |
Subsidiarity |
Problémy je nutno řešit na co možná nejnižší úrovni organizace řízení zdravotní péče. |
Transparentnosti |
Všechny procesy rozhodování je nutno vykonávat transparentně s možností veřejné kontroly (např. obstarávání diagnostické techniky a přístrojového vybavení pro molekulárně–genetickou laboratoř). |
Opatrnosti |
V případě nepřiměřeně vysokého rizika pro pacienta je nutno pozdržet se zamýšleného konání (např. zavádění nových terapeutických metod apod.). |
Rok |
Organizace |
Název |
1948 |
WMA |
Ženevská deklarace |
1949 |
WMA |
Mezinárodní kodex lékařské etiky |
1964 |
WMA |
Helsinská deklarace práv pacienta |
1997 |
UNESCO |
Všeobecná deklarace o lidském genomu a lidských právech |
1997 |
RE |
Mezinárodní dohovor o lidských právech a biomedicíně |
1998 |
Dodatkový protokol o zákazu klonování člověka | |
2003 |
Dodatkový protokol o transplantacích | |
2005 |
Dodatkový protokol o biomedicínském výzkumu | |
2008 |
Dodatkový protokol o genetickém testování v medicíně |
Zákon č. |
Název |
1/1993 Sb. |
Ústava České republiky |
101/2000 Sb. |
Zákon o ochraně osobních údajů a o změně některých zákonů |
372/2011 Sb. |
Zákon o zdravotních službách a podmínkách jejich poskytování |
373/2011 Sb. |
Zákon o specifických zdravotních službách |
296/2008 Sb. |
Zákon o zajištění jakosti a bezpečnosti lidských tkání a buněk určených k použití u člověka a o změně souvisejících zákonů (zákon o lidských tkáních a buňkách) |
285/2002 Sb. |
Zákon o darování, odběrech a transplantacích tkání a orgánů a o změně některých zákonů (transplantační zákon) |
Hlava |
Díl |
Paragraf |
Oblast |
II |
1 |
3-12 |
Asistovaná reprodukce |
2 |
12-16 |
Sterilizace | |
3 |
17-20 |
Terapeutická kastrace, testikulární pulpektomie | |
4 |
21-23 |
Změna pohlaví transsexuálních pacientů | |
6 |
28-30 |
Genetické vyšetření | |
III |
- |
33-40 |
Ověřování nových postupů použitím metody, která dosud nebyla v klinické praxi na živém člověku zavedena |
9 Seznam klíčových slov
A
Pojem |
Kapitola |
A |
|
aberace chromosomové |
5.1 Klasifikace mutací |
aberace chromatidové |
5.1 Klasifikace mutací |
adenin |
2.2 Nukleové kyseliny |
adenylátcykláza |
3.4 Regulace genové exprese |
aditivita |
6.2 Kvantitativní znaky v populaci |
AFM |
8.1 Nanotechnologie |
alela |
4.3 Segregace a kombinace vloh |
alela dominantní |
4.3 Segregace a kombinace vloh |
alela recesivní |
4.3 Segregace a kombinace vloh |
alely identické původem |
7.1 Genealogické vyšetření |
alkaptonurie |
7.3 Biochemická genetika |
allozygotní genotyp |
7.1 Genealogické vyšetření |
alogamická populace |
6 Populační genetika |
aminoacyl-tRNA |
3.3 Translace |
aminoacyl-tRNA-syntetáza |
3.3 Translace |
amitóza |
4. Vztah genetické informace k rozmnožování |
anafáze |
4.1 Mitóza a meióza eukaryontních buněk |
aneuploidie |
5.1 Klasifikace mutací |
anortoploidie |
5.1 Klasifikace mutací |
antimediátorová RNA |
3.4 Regulace genové exprexe |
Archibald Garrod |
7.3 Biochemická genetika |
Ashanti De Silva |
8.2 Molekulární medicína |
autogamická populace |
6 Populační genetika |
autozomálně dominantní dědičnost |
7.1 Genealogické vyšetření |
autozomálně recesivně podmíněná hluchoněmota |
7.1 Genealogické vyšetření |
autozomálně recesivní dědičnost |
7.1 Genealogické vyšetření |
autozomy |
2.1 Chromosomy |
autozygotní genotyp |
7.1 Genealogické vyšetření |
B
Pojem |
Kapitola |
B |
|
Barrovo tělísko |
4.5 Genetická informace a pohlaví |
Batesonovo číslo „c“ |
4.4. Vazba genů |
bezpečnost osobních dat |
8.3 Etické problémy |
binární dělení |
4.2 Rozmnožování prokaryontních buněk |
bioinformační makromolekuly |
2. Genetická informace
2.2 Nukleové kyseliny |
biologické mutageny |
5.2 Rozdělení mutagenů |
Bloomův syndrom |
5.3 Reparační mechanismy |
BRCA1 |
viz karcinom prsu |
BRCA2 |
viz karcinom prsu |
C
Pojem |
Kapitola |
C |
|
cAMP |
3.4 Regulace genové exprexe |
CAP |
3.4 Regulace genové exprexe |
coupling |
viz fáze „cis“ |
C-proužkování |
7.2 Cytologická analýza |
cyklin-dependentní kinázy |
4. Vztah genetické informace k rozmnožování |
cykliny |
4. Vztah genetické informace k rozmnožování |
cytogenetika |
7.2 Cytologická analýza |
cytosin |
2.2 Nukleové kyseliny |
D
Pojem |
Kapitola |
D |
|
dědičnost |
2. Genetická informace |
dědičnost křížem |
4.5 Genetická informace a pohlaví |
dědivost |
6.2 Kvantitativní znaky v populaci |
delece |
5.1 Klasifikace mutací |
deplece DNA |
5.1 Klasifikace mutací |
dicentrický chromosom |
5.1 Klasifikace mutací |
Dicer |
3.4 Regulace genové exprexe |
diploidní stav |
4. Vztah genetické informace k rozmnožování |
DNA |
2.2 Nukleové kyseliny |
DNA-čipy |
8.1 Nanotechnologie |
DNA-microarray |
Viz DNA-čipy |
Downův syndrom |
5.1 Klasifikace mutací |
E
Pojem |
Kapitola |
E |
|
Edwardsův syndrom |
5.1 Klasifikace mutací |
efektivní velikost populace |
6 Populační genetika
6.1 Kvalitativní znaky v populaci |
elongace replikace |
3.1 Replikace |
elongace translace |
3.3 Translace |
endomitóza |
4. Vztah genetické informace k rozmnožování
4.1 Mitóza a meióza eukaryontních buněk |
enzymová indukce |
3.4 Regulace genové exprexe |
enzymová represe |
3.4 Regulace genové exprexe |
excizní reparace |
5.3 Reparační mechanismy |
exony |
3.2 Transkripce |
externí řízení kvality |
7.4 Molekulárně genetické metody |
F
Pojem |
Kapitola |
F |
|
faktor V (proakcelerin) |
6.1 Kvalitativní znaky v populaci |
familiární výskyt onemocnění |
7.4 Molekulárně genetické metody |
farmakogenetika |
7.3 Biochemická genetika |
fáze „cis“ |
4.4. Vazba genů |
fáze „trans“ |
4.4. Vazba genů |
fenokopie |
5.2 Rozdělení mutagenů |
fenotyp |
2.3 Koncepce genu |
fenylketonurie |
5.1 Klasifikace mutací
7.1 Genealogické vyšetření |
Filadelfský chromosom |
5.1 Klasifikace mutací |
FISH |
viz fluorescenční hybridizace in situ |
fluorescenční hybridizace in situ |
8.1 Nanotechnologie |
fotoreaktivace |
5.3 Reparační mechanismy |
fyzikální mutageny |
5.2 Rozdělení mutagenů |
G
Pojem |
Kapitola |
G |
|
G1-fáze |
4. Vztah genetické informace k rozmnožování |
G2-fáze |
4. Vztah genetické informace k rozmnožování |
gen |
2.3 Koncepce genu |
genetická mozaika |
7.4 Molekulárně genetické metody |
genetické izoláty |
7.1 Genealogické vyšetření |
genetický drift |
6.1 Kvalitativní znaky v populaci |
genetický polymorfismus |
7.3 Biochemická genetika |
genofond |
6 Populační genetika |
genotyp |
2.3 Koncepce genu |
genotypové frekvence |
6.1 Kvalitativní znaky v populaci |
genotypové určení pohlaví |
4.5 Genetická informace a pohlaví |
genová terapie |
8.2 Molekulární medicína |
genové (alelové) frekvence |
6.1 Kvalitativní znaky v populaci |
genové určení pohlaví |
4.5 Genetická informace a pohlaví |
geny konstitutivní |
3.4 Regulace genové exprexe |
gonozomy |
4.5 Genetická informace a pohlaví |
G-proužkování |
7.2 Cytologická analýza |
guanin |
2.2 Nukleové kyseliny |
H
Pojem |
Kapitola |
H |
|
Hardy-Weinbergův zákon |
6.1 Kvalitativní znaky v populaci |
HCV |
viz virus hepatitidy C |
hemizygotní stav |
4.5 Genetická informace a pohlaví
7.1 Genealogické vyšetření |
hemofilie |
7.1 Genealogické vyšetření |
hemofilie A |
7.1 Genealogické vyšetření |
hemofilie B |
7.1 Genealogické vyšetření
8.2 Molekulární medicína |
hereditární eliptocytóza |
7.1 Genealogické vyšetření |
heritabilita |
viz dědivost |
heritabilita v širším slova smyslu |
6.2 Kvantitativní znaky v populaci |
heritabilita v užším slova smyslu |
6.2 Kvantitativní znaky v populaci |
heteroduplex |
8.1 Nanotechnologie |
heterogametické pohlaví |
4.5 Genetická informace a pohlaví |
heteroplazmie |
5.1 Klasifikace mutací |
heterotypické dělené buňky |
4.1 Mitóza a meióza eukaryontních buněk |
heterozygot |
4.3 Segregace a kombinace vloh |
Hippokratova přísaha |
8.3 Etické problémy |
holandrická dědičnost |
Viz přímá dědičnost |
homeotypické dělení buňky |
4.1 Mitóza a meióza eukaryontních buněk |
homogametické pohlaví |
4.5 Genetická informace a pohlaví |
homoplazmie |
5.1 Klasifikace mutací |
homozygot |
4.3 Segregace a kombinace vloh |
HPV |
7.4 Molekulárně genetické metody |
HPV 16 |
7.4 Molekulárně genetické metody |
HPV 18 |
7.4 Molekulárně genetické metody |
HPV 31 |
7.4 Molekulárně genetické metody |
HPV 33 |
7.4 Molekulárně genetické metody |
HPV 35 |
7.4 Molekulárně genetické metody |
HRT (high resolution techniques) |
7.2 Cytologická analýza |
Huntingtonova chorea |
8.3 Etické problémy |
hybridizace |
8.1 Nanotechnologie |
hypofosfatémie |
7.1 Genealogické vyšetření |
CH
Pojem |
Kapitola |
Ch |
|
Chargaffova pravidla |
2.2 Nukleové kyseliny |
chemické mutageny |
5.2 Rozdělení mutagenů |
chí-kvadrátový test |
6.1 Kvalitativní znaky v populaci |
chimérismus |
5.1 Klasifikace mutací |
chromosomové určení pohlaví |
4.5 Genetická informace a pohlaví |
chromosomy |
2.1 Chromosomy |
chromozomální aberace |
5.1 Klasifikace mutací |
I
Pojem |
Kapitola |
I |
|
IBD |
viz alely identické původem |
inbrední koeficient |
7.1 Genealogické vyšetření |
inbríding |
7.1 Genealogické vyšetření |
informovaný souhlas pacienta |
8.3 Etické problémy |
iniciace replikace |
3.1 Replikace |
iniciace translace |
3.3 Translace |
iniciační kodon |
3.3 Translace |
integrita jedince |
8.3 Etické problémy |
interfáze |
4. Vztah genetické informace k rozmnožování |
interní řízení kvality |
7.4 Molekulárně genetické metody |
introny |
3.2 Transkripce |
inverze |
5.1 Klasifikace mutací |
inverze paracentrická |
5.1 Klasifikace mutací |
inverze pericentrická |
5.1 Klasifikace mutací |
inzerce |
5.1 Klasifikace mutací |
ionizující záření |
5.2 Rozdělení mutagenů |
J
Pojem |
Kapitola |
J |
|
Jesse Gelsinger |
8.2 Molekulární medicína |
K
Pojem |
Kapitola |
K |
|
karcinom děložního čípku |
7.4 Molekulárně genetické metody |
karcinom prsu |
7.4 Molekulárně genetické metody |
katabolická represe |
3.4 Regulace genové exprexe |
Klinefelterův syndrom |
5.1 Klasifikace mutací |
kód genetický |
3.3 Translace |
kodogenní řetězec |
3.2 Transkripce |
koinfekce |
viz virus lidského papilomu |
kontinuální proměnlivost |
6.2 Kvantitativní znaky v populaci |
L
Pojem |
Kapitola |
L |
|
Leidenská mutace faktoru V |
6.1 Kvalitativní znaky v populaci
7.4 Molekulárně genetické metody |
leukocyty |
7.2 Cytologická analýza |
lokus |
2.3 Koncepce genu |
lyonizace |
4.5 Genetická informace a pohlaví |
M
Pojem |
Kapitola |
M |
|
maskulinní a feminní faktory |
4.5 Genetická informace a pohlaví |
maternální fenylketonurie |
5.1 Klasifikace mutací |
matroklinita |
viz mitochondriální dědičnost |
meióza |
4. Vztah genetické informace k rozmnožování
4.1 Mitóza a meióza eukaryontních buněk |
Mendelova pravidla |
4.3 Segregace a kombinace vloh |
metabolická dráha |
7.3 Biochemická genetika |
metafáze |
4.1 Mitóza a meióza eukaryontních buněk |
metyléntetrahydrofolát reduktáza |
7.3 Biochemická genetika |
migrace |
6.1 Kvalitativní znaky v populaci |
mikroskopie atomárních sil |
8.1 Nanotechnologie |
miRNA |
3.4 Regulace genové exprexe |
mitochondriální dědičnost |
7.1 Genealogické vyšetření |
mitochondrie |
2.1 Chromosomy |
mitotické jedy |
7.2 Cytologická analýza |
mitóza |
4. Vztah genetické informace k rozmnožování
4.1 Mitóza a meióza eukaryontních buněk |
model operonový |
3.4 Regulace genové exprexe |
monozomie |
5.1 Klasifikace mutací |
Moragnova pravidla |
4.4. Vazba genů |
Morganovo číslo „p“ |
4.4. Vazba genů |
mozaicismus |
5.1 Klasifikace mutací |
mRNA (mediátorová RNA) |
2.2 Nukleové kyseliny |
mtDNA depleční syndrom |
5.1 Klasifikace mutací |
mutabilita |
5 Mutace a poškození genetické informace |
mutace |
5 Mutace a poškození genetické informace
6.1 Kvalitativní znaky v populaci |
mutace bodová |
5.1 Klasifikace mutací |
mutace gametické |
5.1 Klasifikace mutací |
mutace genomové |
5.1 Klasifikace mutací |
mutace genové |
5.1 Klasifikace mutací |
mutace chromosomové |
5.1 Klasifikace mutací |
mutace indukované |
5.2 Rozdělení mutagenů |
mutace letální |
5.1 Klasifikace mutací |
mutace neutrální |
5.1 Klasifikace mutací |
mutace přímá |
5.1 Klasifikace mutací |
mutace somatické |
5.1 Klasifikace mutací |
mutace spontánní |
5.2 Rozdělení mutagenů |
mutace tichá |
5.1 Klasifikace mutací |
mutace vitální |
5.1 Klasifikace mutací |
mutace zpětná |
5.1 Klasifikace mutací |
mutageny |
5.2 Rozdělení mutagenů |
N
Pojem |
Kapitola |
N |
|
negativní kontrola |
7.4 Molekulárně genetické metody |
nondisjunkce chromatid |
5.1 Klasifikace mutací |
NOR-proužkování |
7.2 Cytologická analýza |
nukleozom |
2.1 Chromosomy |
O
Pojem |
Kapitola |
O |
|
Okazakiho fragmenty |
3.1 Replikace |
operon |
3.4 Regulace genové exprexe |
ortoploidie |
5.1 Klasifikace mutací |
osteomalácie |
7.1 Genealogické vyšetření |
P
Pojem |
Kapitola |
P |
|
panmixie |
6 Populační genetika |
Pataův syndrom |
5.1 Klasifikace mutací |
PCR |
viz polymerázová řetězová reakce |
polyadenylace |
3.2 Transkripce |
polymerázová řetězová reakce (PCR) |
7.4 Molekulárně genetické metody |
polyploidie |
5.1 Klasifikace mutací |
pozitivní kontrola |
7.4 Molekulárně genetické metody |
pravidlo o čistotě gamet |
4.3 Segregace a kombinace vloh |
pravidlo o nezávislé kombinaci genů |
4.3 Segregace a kombinace vloh |
pravidlo štěpení znaků |
4.3 Segregace a kombinace vloh |
pravidlo uniformity a reciprocity |
4.3 Segregace a kombinace vloh |
premutagen |
5.2 Rozdělení mutagenů |
primární poměr pohlaví |
4.5 Genetická informace a pohlaví |
princip autonomie |
8.3 Etické problémy |
princip non-maleficience |
8.3 Etické problémy |
princip beneficience |
8.3 Etické problémy |
princip opatrnosti |
8.3 Etické problémy |
princip solidarity |
8.3 Etické problémy |
princip spravelnosti |
8.3 Etické problémy |
princip subsidiarity |
8.3 Etické problémy |
princip transparentnosti |
8.3 Etické problémy |
proband |
7.1 Genealogické vyšetření |
profáze |
4.1 Mitóza a meióza eukaryontních buněk |
proměnlivost |
2. Genetická informace |
promotor |
3.2 Transkripce |
prstencové (ring) chromosomy |
5.1 Klasifikace mutací |
příbuzenské křížení |
7.1 Genealogické vyšetření |
přímá dědičnost |
4.5 Genetická informace a pohlaví |
Q-proužkování |
7.2 Cytologická analýza |
R
Pojem |
Kapitola |
R |
|
rachitida |
7.1 Genealogické vyšetření |
Recklinghausenova nemoc |
viz segmentární neurofibromatóza |
rekombinace |
4.1 Mitóza a meióza eukaryontních buněk |
reparace zlomu |
5.1 Klasifikace mutací |
replikace semikonzervativní |
3.1 Replikace |
repulsion |
viz fáze „trans“ |
restituce zlomu |
5.1 Klasifikace mutací |
ribozomy |
3.3 Translace |
RISC |
3.4 Regulace genové exprexe |
RNA |
2.2 Nukleové kyseliny |
RNA interference |
3.4 Regulace genové exprexe |
RNA-polymeráza |
3.2 Transkripce |
rodokmen |
7.1 Genealogické vyšetření |
R-proužkování |
7.2 Cytologická analýza |
rRNA |
2.2 Nukleové kyseliny |
řetězec opožďující se |
3.1 Replikace |
řetězec vedoucí |
3.1 Replikace |
S
Pojem |
Kapitola |
S |
|
Segmentární neurofibromatóza |
7.1 Genealogické vyšetření |
sekundární poměr pohlaví |
4.5 Genetická informace a pohlaví |
selekce |
6.1 Kvalitativní znaky v populaci |
semikvantitativní znaky |
6.2 Kvantitativní znaky v populaci |
sestřih |
3.2 Transkripce |
S-fáze |
4. Vztah genetické informace k rozmnožování |
siRNA |
3.4 Regulace genové exprexe |
sociální segregace |
7.1 Genealogické vyšetření |
solenoid |
2.1 Chromosomy |
sonda |
8.1 Nanotechnologie |
spolehlivost získaných výsledků |
7.4 Molekulárně genetické metody |
srpkovitá anémie |
5.1 Klasifikace mutací
6.1 Kvalitativní znaky v populaci |
SSCP |
7.4 Molekulárně genetické metody |
standardní operační postupy |
7.4 Molekulárně genetické metody |
startovací nukleotid |
3.2 Transkripce |
stupeň konsangvinity |
7.1 Genealogické vyšetření |
substituce |
5.1 Klasifikace mutací |
superfemale |
5.1 Klasifikace mutací |
supermale |
5.1 Klasifikace mutací |
syndrom Cri du chat |
5.1 Klasifikace mutací |
T
Pojem |
Kapitola |
T |
|
Taq-polymeráza |
7.4 Molekulárně genetické metody |
telofáze |
4.1 Mitóza a meióza eukaryontních buněk |
terciární poměr pohlaví |
4.5 Genetická informace a pohlaví |
terminace replikace |
3.1 Replikace |
terminace translace |
3.3 Translace |
terminátor |
3.2 Transkripce |
Thermophilus aquaticus |
7.4 Molekulárně genetické metody |
thymin |
2.2 Nukleové kyseliny |
tolerantní reparace |
5.3 Reparační mechanismy |
transkripční faktory |
3.2 Transkripce |
transkripční jednotky |
3.2 Transkripce |
translační faktory |
3.3 Translace |
translokace nereciproké |
5.1 Klasifikace mutací |
translokace reciproké |
5.1 Klasifikace mutací |
translokace Robertsonské |
5.1 Klasifikace mutací |
trizomie |
5.1 Klasifikace mutací |
tRNA |
2.2 Nukleové kyseliny |
Turnerův syndrom |
5.1 Klasifikace mutací |
Typ Abraxas |
Viz chromosomové určení pohlaví |
Typ Drosophila |
Viz chromosomové určení pohlaví;
Viz genové určení pohlaví |
Typ Habrobracon |
Viz chromosomové určení pohlaví;
Viz genové určení pohlaví |
Typ Lymantria |
Viz genové určení pohlaví |
U
Pojem |
Kapitola |
U |
|
úpravy kotranslační |
3.3 Translace |
úpravy posttranslační |
3.3 Translace |
uracyl |
2.2 Nukleové kyseliny |
Ústava České republiky |
8.3 Etické problémy |
ústřední dogma molekulární biologie |
3 Zpracování genetické informace |
UV záření |
5.2 Rozdělení mutagenů |
V
Pojem |
Kapitola |
V |
|
vazba 3´-5´ fosfodiesterická |
2.2 Nukleové kyseliny |
vazba N-glykosidická |
2.2 Nukleové kyseliny |
vektor |
8.2 Molekulární medicína |
virus hepatitidy typu C |
7.4 Molekulárně genetické metody |
virus lidského papilomu |
7.4 Molekulárně genetické metody |
vitamin D rezistentní rachitida |
7.1 Genealogické vyšetření |
vrozená vývojová vada |
5.2 Rozdělení mutagenů |
W
Pojem |
Kapitola |
W |
|
Weber-Cockayneov syndrom |
5.3 Reparační mechanismy |
Wolfův-Hirschhornův syndrom |
5.1 Klasifikace mutací |
X
Pojem |
Kapitola |
X |
|
X – vázaná dominantní dědičnost |
7.1 Genealogické vyšetření |
X – vázaná recesivní dědičnost |
7.1 Genealogické vyšetření |
Xeroderma pigmentosum |
5.3 Reparační mechanismy |
Z
Pojem |
Kapitola |
Z |
|
Zákon č. 373/2011 Sb., o specifických zdravotních službách |
8.3 Etické problémy |
zesilovač transkripce |
3.4 Regulace genové exprexe |
zeslabovač transkripce |
3.4 Regulace genové exprexe |
zlom |
5.1 Klasifikace mutací |
znaky neúplně vázané na pohlaví |
4.5 Genetická informace a pohlaví |
znaky pohlavně ovládané |
4.5 Genetická informace a pohlaví |
znaky úplně vázané na pohlaví |
4.5 Genetická informace a pohlaví |
zpětná transkripce |
3.2 Transkripce |
životní cyklus buňky |
4. Vztah genetické informace k rozmnožování |