Genetika
2.2 Nukleové kyseliny
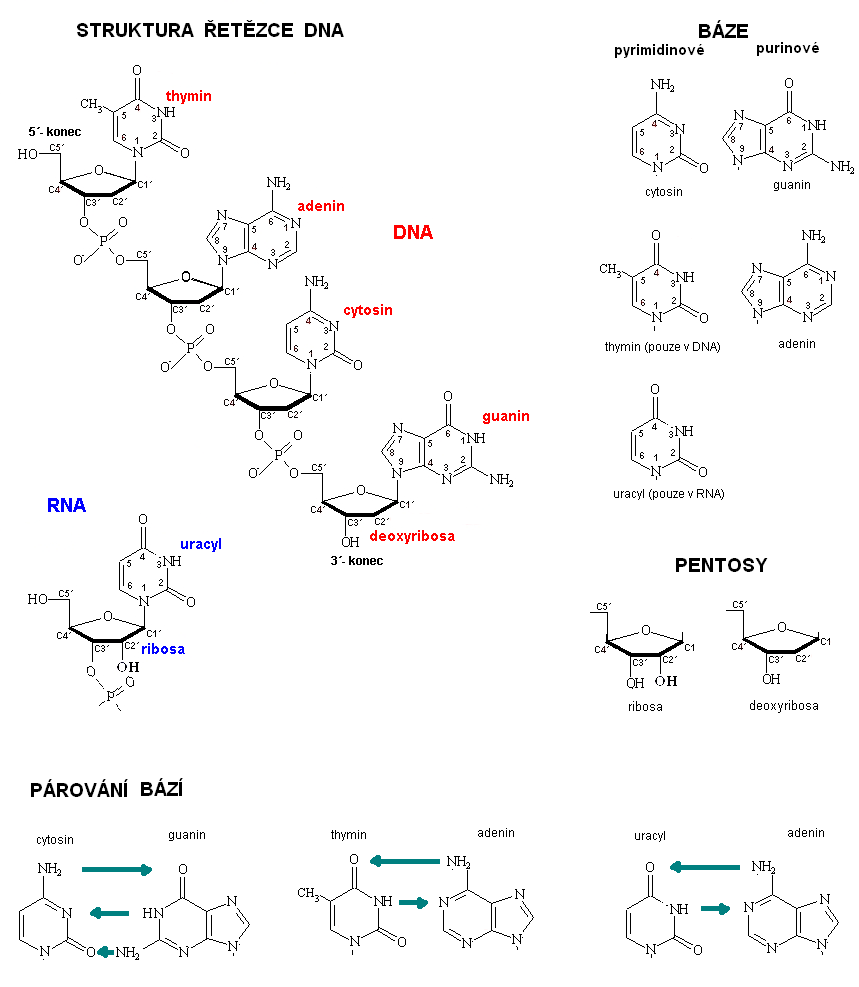
Mezi bioinformační makromolekuly, které nazýváme nukleové kyseliny, patří DNA a RNA. Název DNA je odvozen z anglického (deoxyribonucleic acid), protože obsahuje β -D-2-deoxyribosu. Je to cukr s pěti atomy uhlíku (cyklická furanosa).
Kromě pentosy zde nacházíme i dusíkaté báze. Jsou to heterocyklické sloučeniny, kde je heteroatomem dusík (obrázek 1). Jsou to deriváty purinu nebo pyrimidinu. V DNA se nacházejí dvě purinové báze - adenin a guanin. Pyrimidinové báze jsou tři - cytosin, thymin a uracyl.
Dusíkaté báze se vážou svým heteroatomem dusíku na C1' uhlík pentosy za vzniku N-glykosidické vazby. Pyrimidinové báze se vážou prostřednictvím dusíku N1, purinové bázi se vážou dusíkem N9. Spojením dusíkaté báze s pentosou prostřednictvím N-glykosidické vazby vzniká v případě DNA tzv. deoxyribonukleozid.
Vytvořením esterové vazby mezi zbytkem kyseliny fosforečné a C5' atomem uhlíku pentosy v deoxyribonukleozidu vzniká deoxyribonukleotid-5'-monofosfát. Zbytek kyseliny fosforečné se může vázat další esterovou vazbou na C3' atom uhlíku jiného deoxyribonukleotidu. Vzniká tak 3 ', 5'-fosfodiesterická vazba, která spojuje uhlík C5' jednoho deoxyribonukleotidu s atomem C3' následujícího deoxyribonukleotidu (obrázek 1).
Biologickou funkci nosiče genetické informace má však až dvouřetězcová DNA (dále dsDNA). Její strukturu poprvé popsali v roce 1953 J. Watson, F. Crick a M. Wilkins (Nobelova cena za fyziologii a medicínu v roce 1962). Na objevu se výrazným způsobem podílela i Rosaline Franklin. dsDNA se skládá ze dvou polydeoxyribonukleotidových řetězců, které jsou spojeny vodíkovými vazbami. Ty vznikají vzájemným párováním dusíkatých bází tak, že adenin se dvěma vodíkovými vazbami spojí s thyminem a cytosin se třemi vodíkovými vazbami váže s guaninem (A=T; C≡G; viz obrázek 1).
Vzájemný poměr bázi, kterými se párují oba řetězce se řídí Chargaffovými pravidly:
- Molární obsah adeninu se rovná molárnímu obsahu thyminu:
A = T; T = A
- Molární obsah guaninu se rovná molárnímu obsahu cytosinu:
G = C; C = G
- Obecně je v každé dvoušroubovici DNA obsah molekul purinových bází rovný obsahu molekul pyrimidínových bází:
(A + G) = (C + T)
- Molární obsah adeninu se rovná molárnímu obsahu thyminu:
A = T; T = A
- Molární obsah guaninu se rovná molárnímu obsahu cytosinu:
G = C; C = G
- Obecně je v každé dvoušroubovici DNA obsah molekul purinových bází rovný obsahu molekul pyrimidínových bází:
(A + G) = (C + T)
Příklad: Sekvenci dvoušroubovice DNA tvoří 10 bází:
5´ – ATTAGCCATA – 3´
3´ – TAATCGGTAT – 5´
7 A = 7 T
3 C = 3 G
10 (C + T) = 10 (A + G)
Na uvedeném příkladu můžeme vidět tzv. komplementaritu obou řetězců DNA, která je daná párováním bází A-T a C-G. Důležitou vlastností dsDNA je antiparalelní orientace obou řetězců, z nichž je vytvořena. Můžeme totiž určovat tzv. orientaci řetězců, které na jednom konci končí deoxyribosou, na jejíž C5' uhlíku již není estericky vázán zbytek kyseliny fosforečné. Opačný konec je tvořen deoxyribosou, která má volnou OH skupinu na 3' uhlíku místo toho, aby byla esterická vazba s 5' uhlíkem deoxyribosy dalšího deoxyribonukleotidu (obrázek 1). Antiparalelní orientace je tedy stav, kdy se vzájemně spojí dva řetězce DNA s pomocí komplementárního párování bází, přičemž jejich směr je opačný. 5'- konec jednoho řetězce sousedí s 3' -koncem protějšího komplementárního řetězce a naopak.
U DNA známe několik úrovní strukturální organizace molekuly. Primární strukturou DNA se rozumí pořadí, čili sekvence deoxyribonukleotidů v řetězci DNA. Sekundární strukturou je samotná dvoušroubovice, tvořena dvěma antiparalelně orientovanými řetězci, které jsou spojeny vodíkovými vazbami. Ty vznikají při párování bází ve smyslu Chargaffových pravidel. Terciární struktura DNA vzniká při spiralizaci dsDNA, přičemž vzniká tzv. superhelix, čili nadšroubovice. Ta je důležitá v tvorbě struktur chromozomů během buněčného dělení. Enzymy v těchto dějích se nazývají topoisomerasy.
RNA je na rozdíl od DNA tvořena pouze jedním polyribonukleotidovým řetězcem. Druhým zásadním rozdílem je typ pentosy. RNA obsahuje β -D-ribosu, která má na C2' -uhlíku OH skupinu. Třetím rozdílem RNA od DNA je obsah uracylu místo thyminu. Rozdíly mezi oběma typy nukleových kyselin přehledně uvádí tabulka 1.
Tabulka 1/A Rozdíly ve složení nukleových kyselin
Typ nukleové kyseliny |
Stavební složky | ||
Pentosa |
Dusíkaté bázy |
Fosfodiesterická vazba | |
DNA |
β-D-2-deoxyribosa |
adenin, thymin, cytosin, guanin |
estericky vázaný zbytek H3PO4 |
RNA |
β -D-ribosa |
adenin, uracyl, cytosin, guanin |
estericky vázaný zbytek H3PO4 |
Známe několik typů RNA:
- mRNA (mediátorová RNA, angl. messenger RNA) přenáší informaci o pořadí aminokyselin z jádra buňky do cytoplasmy.
- rRNA (ribozomální RNA) je součástí struktury obou podjednotek, z nichž jsou tvořeny ribozomy. Podílí se na strukturální architektuře ribozomu a hraje důležitou roli v procesu syntézy bílkovin, tedy proteosyntéze.
- tRNA (transferová RNA) slouží pro přenos aktivovaných aminokyselin k ribozomům, na kterých se děje syntéza nového polypeptidového řetězce. tRNA je tvořena přibližně 70-80 ribonukleotidy. Přestože je tvořena jediným řetězcem RNA, jeho některé části obsahují vzájemně komplementární uspořádání bází. Důsledkem toho dochází k intramolekulárnímu párování bází, přičemž vzniká typická sekundární struktura, která svým vzhledem připomíná jetelový list. Jeho "listy" tvoří tři ramena: antikodonové, dihydrouridinové a pseudouridinové. Na opačném konci je tzv. akceptorové rameno.
Antikodonové rameno má důležitou roli při překladu genetické informace (genetického kódu) z pořadí nukleotidů v nukleových kyselinách do pořadí aminokyselin v polypeptidech během translace. Na akceptorovém rameni je navázána příslušná aktivovaná aminokyselina. Zvláštní název pseudouridinového a dihydrouridinového ramene je odvozen od obsahu atypických, tzv. minoritních bází pseudouridinu a dihydrouridinu.
Interakcemi mezi rameny, jejichž podstatou je především tvorba vodíkových můstků, vzniká terciární struktura RNA.
Obrázek 1 - Nukleové kyseliny