Hematologie
3.1.2 Hemoglobin
3.1.2 Hemoglobin
Hemoglobin je červené krevní barvivo a je jednou z nejdůležitějších složek erytrocytů. Jeho koncentrace u mužů je 135–170 g Hb/l krve, u žen 120–158 g/l krve. Jeho hlavní úlohou je vazba a přenos kyslíku v těle, uplatňuje se také v transportu oxidu uhličitého. Při plném nasycení váže 1 g hemoglobinu 1,34 ml kyslíku, celkový objem krve může tedy vázat okolo 1 litru kyslíku.
Molekula hemoglobinu je tetramer, skládá se tedy ze 4 podjednotek, kde každou podjednotku tvoří bílkovinná složka (polypeptidový řetězec) – globin a prostetická skupina – hem, na který se v monomeru globinů váže molekula kyslíku. Za červenou barvu krve je zodpovědný právě hem díky svému systému konjugovaných dvojných vazeb. Vždy dvě a dvě podjednotky hemoglobinu jsou stejné.
Základem struktury hemu jsou cyklicky vázané 4 pyrolové kruhy spojené methinovými můstky, které vytváří protoporfyrin IX. Uprostřed tetrapyrolového skeletu je koordinačně kovalentními vazbami vázáno dvojmocné železo (Fe2+), které i po vazbě kyslíku zůstává dvojmocné (Obrázek 3.1). Železo v hemu je šestivazné (4 vazby s atomy dusíku pyrolových kruhů, 1 vazba s histidinem v globinu a poslední je vazba s O2).
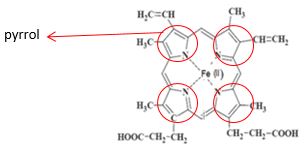
Obrázek 3.1: Hem – protoporfyrin IX
Zdroj: autor
Globin je syntetizován na ribozomech, zatímco syntéza hemu probíhá převážně v mitochondriích a je závislá na množství železa dodávaného do erytroidních buněk. Hlavní úlohu v regulaci syntézy hemoglobinu má hem. Hem stimuluje tvorbu globinu, ovlivňuje přísun železa do buňky a kontroluje vlastní syntézu. Nahromadění hemu tlumí transport železa do buňky a inhibuje aktivitu enzymů katalyzujících syntézu hemu.
Existuje velké množství hemoglobinů, všechny mají stejný hem, ale liší se v globinové části, která určuje typ hemoglobinu. V lidských hemoglobinech se vyskytují polypeptidové řetězce α, β, γ, δ, ε a ζ, které se liší počtem a pořadím aminokyselin. U dospělých převládá (z 98 %) hemoglobin A - HbA (také HbA1; α2β2), který je tvořený dvěma řetězci α a dvěma řetězci β. Minoritní hemoglobin HbA2 (α2γ2) má místo řetězce β, řetězec γ. U plodu se v průběhu nitroděložního života vyskytují embryonální hemoglobin (α2ε2, prvních 10 týdnů) a fetální hemoglobin HbF (α2δ2)), který je po narození nahrazen HbA. Hemoglobin HbF má vyšší afinitu ke kyslíku než hemoglobin dospělých, umožňuje tak snadnější přenos kyslíku z krve matky. V důsledku strukturních mutací genů hemoglobinu vznikají různé poruchy (hemoglinopatie), kdy dochází například k tvorbě srpků - HbS (srpkovitá anémie), k poruše syntézy řetězců (αβ) globinu (thalasemie αβ), ztrátě hemu či disociaci tetrameru. Působením oxidačních činidel (dusičnany, dusitany) může také dojít ke změně afinity ke kyslíku a ke změně v oxidačním čísle železa (z dvojmocného na trojmocné) za vzniku methemoglobinu – HbM (methemoglobinemie). V erytrocytech se běžně tvoří malé množství methemoglobinu, který je systémem methemoglobinreduktáz neustále redukován.
Jedna molekula hemoglobinu může vázat čtyři molekuly kyslíku. Vazba jedné molekuly kyslíku ovlivní vazbu dalších molekul O2. Navázáním první molekuly O2 se zvýší afinita Hb pro další molekuly O2. Poslední, čtvrtý kyslík se váže nejjednodušeji. Vazbou kyslíku na hemoglobin vzniká oxyhemoglobin.
Afinita hemoglobinu ke kyslíku je ovlivněna několika faktory: pH, parciální tlak CO2 a teplota. Zvýšená afinita hemoglobinu ke kyslíku je dána zvýšením pH, snížením pCO2 či snížením teploty. Naopak snížená afinita ke kyslíku je způsobena snížením pH, zvýšením pCO2 či zvýšením teploty. Snížená afinita ke kyslíku usnadňuje uvolnění kyslíku z oxyhemoglobinu. V erytrocytech je afinita hemoglobinu ke kyslíku regulována koncentrací D-2,3-bisfosfoglycerátu (2,3-BPG), jednoho z produktů glykolýzy. Zvýšená produkce 2,3-BPG snižuje afinitu hemoglobinu ke kyslíku (váže se do centra deoxyhemoglobinu, udržuje Hb v deoxy stavu a snižuje tím schopnost hemoglobinu vázat kyslík), zatímco jeho pokles má opačný efekt. Vazba 2,3-BPG na oxyhemoglobin je slabá.
Hemoglobin může vázat také oxid uhličitý za vzniku karbaminohemoglobinu a podílí se tak na jeho transportu krví. Oxid uhličitý se neváže na hem, ale na globinové řetězce Hb. Při oxygenaci se CO2 uvolňuje. Oxid uhličitý se v krvi vyskytuje nejčastěji ve 3 formách:
I. jako hydrogenuličitanový anion
II. rozpuštěný v plazmě
III. navázaný na hemoglobin
Většina CO2 vzniklého v tkáních je do plic přenesena ve formě HCO3-, vzniklého disociací kyseliny uhličité. Vytvořený HCO3- volně proniká membránou erytrocytů do plazmy výměnou za chloridové ionty tzv. chloridový posun neboli Hamburgerův efekt, který je důležitý pro zachování rovnovážného stavu s okolním prostředím (plazmou). Membrána erytrocytů je neprostupná pro kationty a tak je tato výměna důležitá též i pro elektroneutralitu membrány erytrocytů. Koncentrace oxidu uhličitého a chloridů ovlivňují afinitu Hb ke kyslíku. Zvýšená koncentrace CO2 v kapilárách podporuje uvolnění kyslíku z Hb.
Na dvojmocné železo hemu se může vázat také oxid uhelnatý za vzniku karboxyhemoglobinu. Karboxyhemoglobin nemůže vázat kyslík, oxid uhelnatý tak brání v přenosu O2. Vysoká afinita hemoglobinu k oxidu uhelnatému způsobuje, že stačí malé množství CO ve vdechovaném vzduchu, aby došlo ke smrtelné otravě (0,1 % CO ve vzduchu způsobí nasycení poloviny veškerého hemoglobinu v těle). Vazba hemoglobinu s CO je velmi pevná a rozpadá se pomaleji než vazba s kyslíkem.
Neenzymovou reakcí glukózy s řetězci globinů vzniká glykovaný hemoglobin, ten vzniká při dlouhodobě vysoké glykémii. Glukóza zůstává navázána po celou dobu života erytrocytu.