Genetika
7.3 Biochemická genetika
Na počátku 20 století si britský lékař Archibald Garrod všiml společné symptomatologie u pacientů, kteří trpěli alkaptonurií. Tuto poruchu způsobují mutace genu pro enzym homogentizát 1,2-dioxygenáza (HGD). Pokud se v játrech tento enzym následkem mutací inaktivuje, v organismu se hromadí jeden z metabolitů aminokyseliny tyrosinu - kyselina homogentisová. Vylučuje se močí, která proto ztmavne při stání. Polymer kyseliny homogentisové se jako pigment ukládá v chrupavkách a pojivových tkáních, které tím získávají typickou pigmentaci (ochronózu). Také způsobuje artritidu, přičemž nejvíce bývají postiženy velké klouby a páteř.
Garrod si jako první všiml, že uvedený soubor symptomů je přenášen v rodokmenu postižených probandů. Jinými slovy, jedná se o genetickou vlohu. Vzhledem k tomu, že Mendelovy zákony byly v té době právě "znovuobjeveny" a širší odborné veřejnosti stále v podstatě neznámé, podobně jako biochemická podstata metabolických dějů člověka, také Garrodův objev zůstal na několik desetiletí nedoceněný.
Dnes víme, že alkaptonurie je autozomálně recesivní dědičné onemocnění, jehož podstatou je ztráta aktivity enzymu homogentizát 1,2 dioxygenázy. Její gen se nachází u člověka na delším raménku chromozomu č. 3, lokace 3q21-23. Defektní enzym mohou způsobit mutace na více místech nukleotidové sekvence genu. Dnes jich je známých několik desítek. Většina mutantních (recesivních) alel je skryta u heterozygotů, kteří jsou metabolicky zdraví, protože na druhém chromozomu mají standardní "zdravou" alelu. Pokud se u jedince setkají dvě mutantní alely (které nemusí být identické), jedinec je postižený alkaptonurií. V naší populaci je frekvence tohoto onemocnění přibližně 1: 20000 novorozenců.
Příklad alkaptonurie jsme neuvedli náhodou a chceme jím zdůraznit provázanost biochemických metabolických drah člověka. Tato dědičná choroba je důsledkem selhání jednoho z enzymů téže dráhy, kde v jednom z kroků vzniká mutací i fenylketonurie, kterou jsme se podrobně zabývali v kapitole 5.1 Klasifikace mutací (Příklad 1).
Na obrázku 9 můžeme vidět souvislost metabolické dráhy s genetickou konstitucí pro enzymy, které organismus využívá v jednotlivých krocích biochemické dráhy. Na základě příkladu na obrázku 9 můžeme vyvodit následující důležité závěry:
- biochemické pochody v organismu jsou katalyzovány enzymy, jejichž struktura i funkce je dána geneticky,
- metabolismus se skládá z jednotlivých biochemických drah, které jsou vzájemně propojeny,
- mutace poškozuje v metabolické dráze sice pouze jeden krok, může tím však ovlivnit celý další sled biochemických reakcí.
Pokud již jednou vznikla mutace, její výsledný efekt na produkt postiženého genu závisí od toho, na jakém místě tento gen poškodila. Obecně rozeznáváme poruchy struktury enzymu, které způsobí jeho výslednou dysfunkci a poruchy v regulaci kvantitativní produkce enzymu, které ovlivňují jeho výslednou celkovou aktivitu v organismu.
V této souvislosti zdůrazňujeme pojem genetický polymorfismus, který chápeme jako společný výskyt dvou a více alel sledovaného genu v populaci v takové četnosti, že není pravděpodobné, aby byly výsledkem náhodných mutací. Většinou se jako populační hranice početnosti pro tento typ alel považuje 1 %. Z toho vyplývá, že polymorfismem jsou nám již dobře známé krevní skupiny z kapitoly 4.3. Nejsou však zdaleka jediné. Značná část enzymového arzenálu organismu vykazuje heterogenitu tohoto typu.
Obrázek 9 - Spřažení biochemických reakcí v metabolické dráze
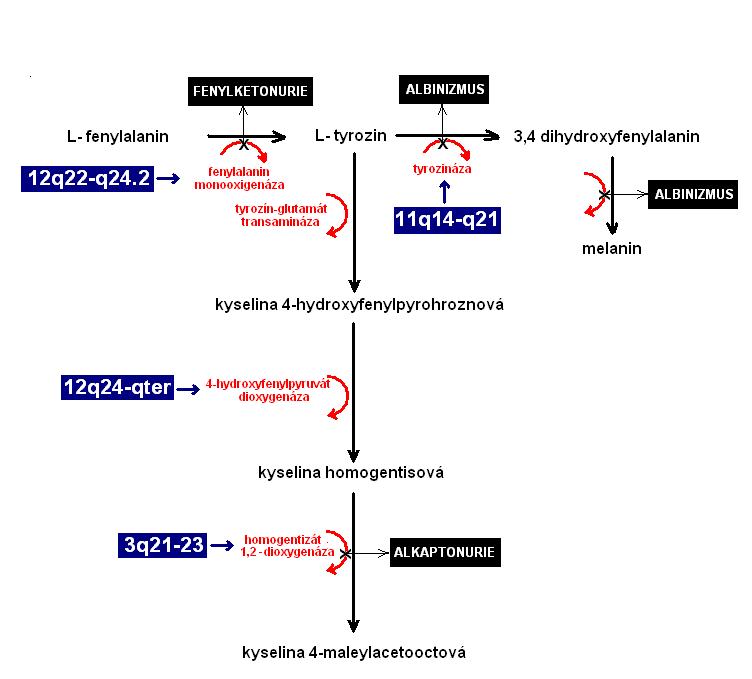
Legenda: enzym fenylalanin monooxigenáza je podmíněná genem, který je lokalizován na delším raménku chromosomu č. 12. Enzym tyrozináza kóduje gen, který se nachází na delším raménku chromosomu č. 11. Poblíž konce delšího raménka chromosomu č. 12 se nachází gen, kódující enzym 4- hydroxyfenylpyruvát dioxigenázu. Struktura a funkce enzymu homogentizát 1,2-dioxigenázy je podmíněná genem, který je lokalizován na delším raménku chromosomu č. 3. Vidíme, že jediná metabolická dráha je podmíněna geny, které jsou umístěny na různých chromozomech.
Výše uvedený učebnicový příklad alkaptonurie a následné vysvětlení genetického polymorfismu má velmi důležitý dopad na využití poznatků ve farmakogenetice. Farmakogenetika v moderním pojetí sleduje genetický individualismus organismu vzhledem na jeho schopnost metabolizace léčebných přípravků. Jako příklad jsme nastínili různou schopnost jedinců rozeznávat hořkou chuť PTC (kapitola 6.2; obrázek 5). Podobná variabilita platí i pro jiné farmaceutické produkty. Je známá např. senzitivita na antimalarikum primaquin v důsledku deficitu aktivity glyceraldehyd-3-fosfátdehydrogenázy, nebo vznik periferní neuropatie při podání isoniazidu v důsledku zpomalené metabolizace (acetylace) tohoto antituberkulotika.