Genetika
8.1 Nanotechnologie
Hlavní trend současného vývoje laboratorních vyšetřovaných metod, ke kterým patří i genetické a molekulárně-genetické testování, se vyvíjí především směrem k automatizaci a miniaturizaci. To je předpokladem pro urychlení analýz a minimalizaci lidského faktoru jako hlavní příčiny hrubých a náhodných chyb.
Miniaturizace laboratorních analýz zároveň vytváří reálný předpoklad pro výrazné zvýšení množství vzorků nebo genů, které jsou vyšetřovány současně. Znamená to, že pokud pátráme po rizikové alele jediného genu, můžeme vyšetřit větší počet pacientů najednou. Pokud chceme naopak vyšetřit u jediného pacienta přítomnost alel několika genů nebo jejich aktivitu, rovněž to můžeme provést současně.
Dalším nezanedbatelným rysem těchto metod je jejich příklon k tzv. "suché fázi". Všichni studenti si jistě pamatují na praktické hodiny v chemické laboratoři. Chemické reakce byly téměř výhradně prováděny ve zkumavkách. Jinými slovy, tyto reakce probíhaly v homogenním, kapalném prostředí. Na této bázi byla postavena nejen "klasická" klinická biochemie, ale také molekulárně-genetická diagnostika (viz kapitoly 7.3 Biochemická genetika a 7.4 Molekulárně genetické metody).
V současnosti již pokrok zašel dál. Dnes se rutinně využívají testovací systémy, které nejsou založeny na chemické reakci v homogenním kapalném prostředí. Příkladem je např. stanovení glykémie s pomocí kapesních analyzátorů. Kapka vzorku se nanese na suchý testovací proužek. Na něm jsou pevně navázány všechny potřebné komponenty (enzymy) a po přidání vzorku se přístrojem stanoví míra změny fyzikálně-chemických parametrů, která je úměrná koncentraci stanovovaného parametru. Podobný rozvoj se udál i v oblasti molekulárně - genetických metod a vyústil do vývoje DNA-čipů (DNA-mikročip, angl. DNA-microarray).
Základem vývoje těchto metod je schopnost dvou oddělených DNA-řetězců (nebo i RNA - řetězců) spárovat se do jediné dvoušroubovicové molekuly DNA. Je to možné tehdy, pokud jsou oba řetězce vzájemně komplementární (viz kapitola 2.2 Nukleové kyseliny). Mezi jednotlivými bázemi se vytvoří vodíkové vazby ve smyslu Chargaffových pravidel: adenin se spojí dvěma vodíkovými vazbami s tyminem a cytosin se spojí třemi vodíkovými vazbami s guaninem. K tomuto ději dochází úplně přirozeně např. v průběhu transkripce. Syntéza RNA se tehdy děje podle jednoho ze dvou řetězců, které se po dokončení procesu znovu spojí do jediné dvouřetězcové molekuly DNA.
K spárování však může dojít i v případě, že oba řetězce jsou sice komplementární, ale liší se svým původem. Pokud uměle vytvoříme (nově syntetizujeme) řetězec DNA, ten se může stejně spolehlivě navázat na druhý řetězec DNA, který pochází z buňky člověka. Jedinou podmínkou pro tento proces je již zmíněná komplementarita řetězců. Proces tvorby dvoušroubovice, která vznikla spárováním dvou řetězců různého původu, nazýváme hybridizace. Samotnou dvoušroubovicovou molekulu DNA, která vznikla tímto způsobem, nazýváme heteroduplex.
V devadesátých letech minulého století došlo k rozvoji prvních hybridizačních technik. Na pevný podklad (např. nylonovou membránu) se ukotvili (např. působením vyšší teploty) řetězce DNA (tzv. sonda). Následně se k nim přidali vzorky, které obsahovaly denaturovanou DNA (tj. s oddělenými řetězci) značenou radioaktivně nebo fluorescenčně. Pokud se ve vzorku DNA nacházela sekvence bází, která byla komplementární se sekvencí navázané DNA-sondy na membráně, vytvořila se dvoušroubovice. Její vznik se ověřil určením přítomnosti signálu na membráně (radioaktivního záření nebo fluorescence).
Uvedený postup je příkladem tzv. Southernovy hybridizace, tj. hybridizace dvou řetězců DNA. Hybridizace řetězců RNA se nazývá northernová hybridizace. Tato klasická, manuální technika dovoluje testovat najednou několik desítek vzorků.
Miniaturizace a robotizace celého procesu vedli k metodikám DNA – microarrays, které jsou využívány v dnešní době. Jejich princip je v podstatě stejný jako u klasické hybridizace (obrázek 13).
Na pevnou podložku se nanese tzv. sonda, kterou tvoří krátké řetězce DNA (tzv. oligomery). Ty obsahují pouze několik desítek nukleotidů. Díky miniaturizaci celé technologie je možné najednou ukotvit až několik desítek tisíc různých sond, které jsou jako pravidelná struktura bodů rovnoměrně rozloženy na skleněném nosiči (obrázek 13). Po nanesení vzorku s neznámou DNA, která je fluorescenčně značená, nastává hybridizace. Ta proběhne pouze tehdy, pokud jsou sekvence nukleotidů řetězců DNA ve vzorku komplementární se sekvencí nukleotidů DNA sondy. Po excitaci světlem o vhodné vlnové délce vyzáří molekula fluoroforu, kterou je vzorek označen, světlo se specifickou barvou (tj. o specifické vlnové délce). Podle intenzity tohoto světla lze určit, zda k hybridizaci došlo, a v jaké míře. Pochopitelně, i tato metodika má ještě nevyřešené problémy. Mezi ně patří zejména:
- Vysoká finanční nákladnost vyšetření, která brání jeho většímu rozšíření v rutinní genetické diagnostice a zůstává doménou specializovaných pracovišť,
- technické nedostatky, jakými jsou poměrně vysoká míra pozadí signálu, která vadí hlavně v případě slabého vyzářeného světla. Důsledkem je nedostatečná senzitivita. Dále jsou to problémy statistického vyhodnocení výsledků.
- Metodologické problémy charakterizuje zejména nedostatečná standardizace postupů a obtížná reproducibilita (opakovatelnost vyšetření).
Naopak, mezi slibné výhody, které metodika poskytuje, patří především:
- Výrazný rozvoj testování genové exprese v rámci farmakoterapie, která by byla přizpůsobena geneticky-metabolickým specifikům pacienta,
- několikanásobné zvýšení kapacity a rychlosti vyšetření, v důsledku čehož se celý proces vyšetření vzorků pacientů výrazně zrychlí,
- komplexnost vyšetření genotypu jedince, důsledkem čeho vzroste diagnostická výpovědní hodnota vyšetření se zpětným vlivem na léčbu a celkovou prognózu pacienta.
Poznámka: hybridizační techniky, které jsou založeny na vzájemné komplementaritě párování bází dvou řetězců DNA různého původu, nejsou doménou pouze molekulární genetiky. V kapitole 7.2 Cytologická analýza jsme se zabývali způsoby vizualizace a vyšetření chromosomů. Významnou podmínkou proužkovacích technik je kultivace buněk ze vzorku pacienta a zastavení jejich dělení v metafázi mitózy. Tento požadavek nevyžaduje metoda FISH (fluorescenční hybridizace in situ, z ang. fluorescence in situ hybridization). V tomto případě se buňky ze vzorku pacienta nemusí zpracovat tak, aby byly viditelné spiralizované chromosomy v metafázi. Po fixaci buněk v interfázi a použití fluorescenčně značené sondy (jednořetězcová DNA) se v případě její vazby na DNA preparátu potvrdí přítomnost sekvence DNA, kterou ve vzorku hledáme. Tímto způsobem se může ověřit např. přítomnost trizómií nebo chromozomálních aberací. FISH má díky tomu značné uplatnění také v reprodukční medicíně, protože v případě asistované reprodukce je množství biologického materiálu ve vzorku značně omezené a neumožňuje provést klasickou cytogenetickou analýzu.
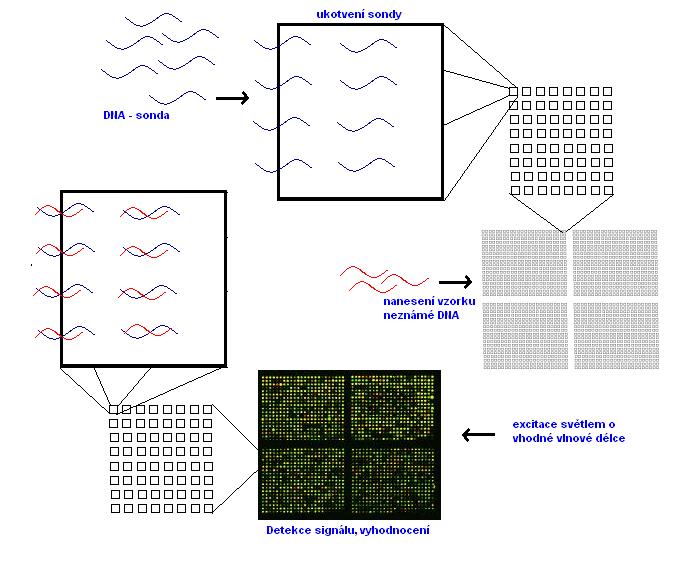
Obrázek 13 - Princip DNA – microarray
Výrazný pokrok lze sledovat také ve vývoji zobrazovacích metod. Mikroskopie atomárních sil (z angl. atomic force microscope, AFM; synonymní název scanning force microscope, SFM) je v současnosti používána a testována v primárním výzkumu v biologii a genetice. Schopností rozlišení struktury vzorku (několik nm) se přibližuje možnostem elektronové mikroskopie a podstatně převyšuje možnosti optického mikroskopu.
Metoda se liší od optického mikroskopu (a také od transmisního elektronového mikroskopu) tím, že zobrazuje pouze povrch fixovaného vzorku (jako např. skenovací elektronová mikroskopie, SEM). Na rozdíl od SEM však nevyžaduje náročnou přípravu vzorek a dokáže pracovat v přirozeném prostředí biologických molekul. Pro DNA nebo RNA je přirozeným prostředím kapalné prostředí. Nukleové kyseliny mají totiž mimo jiné na svém povrchu z molekul vody vytvořenou jakousi "hydratační páteř" (angl. spin of hydration). Proto je studium jejich struktury v přirozeném prostředí velmi důležité.
Principem metody (obr. 14) je využití miniaturního ostrého hrotu, který je upevněn na ohebném nosníku. Druhý konec nosníku je pevně ukotven. Hrot se rovnoměrně pohybuje těsně nad povrchem vzorku (molekuly DNA, proteinu, hormonu apod.). Díky malé vzdálenosti mezi hrotem a vzorkem působí na hrot elektrostatické síly, které v závislosti na konkrétní struktuře rozkmitají ohebný nosník s hrotem. Kmitání, způsobeno atomárními sílami, je zachyceno laserovým paprskem a následně převedeno do trojrozměrného obrazu.
Obrázek 14 - Princip mikroskopie atomárních sil

Metoda je stále ještě ve vývoji a uvedený princip je pouze hrubým náčrtem jejích potenciálních možností. Bude však nutné odstranit nejvážnější nedostatky, kterými jsou malý rozsah velikosti snímaného pole (velikosti obrazu), pomalost snímání a deformity obrazu.
Oba uvedené příklady metod jsme vybrali proto, že už dnes jsou výrazně rozpracovány i po metodické stránce. Z hlediska potenciálu jejich využití v genetice představují slibnou perspektivu. Pochopitelně, existuje mnohem širší spektrum vyvíjených nanotechnologických aplikaci. Jejich rutinní zavedení v genetice je však ještě otázkou dlouhého vývoje a výzkumu.